RESEARCH ARTICLE
Effect of Deferiprone on Hepatic Expression of Hamp, Ftl, and Tfr1 Genes in an Iron-Overloaded Rat (Rattus norvegicus) Model
Academic Editor: Pilli Govindaiah
Sciences of Pharmacy|Vol. 4, Issue 4, pp. 301-305 (2025)
Views
Downloads
Shares
Received
Jun 24, 2025Revised
Jul 31, 2025Accepted
Aug 5, 2025Published
Dec 25, 2025
Abstract
Introduction
Iron is a vital micronutrient involved in numerous physiological processes, including redox reactions, oxygen transport, DNA synthesis, and various enzymatic activities (1). Despite a relatively small daily requirement of approximately 20–30 mg, the human body contains 3–6 grams of iron, predominantly stored in the ferrous (Fe²⁺) and ferric (Fe³⁺) forms (2). However, excess iron can be toxic, catalyzing the formation of reactive oxygen species (ROS) and triggering oxidative stress and cellular damage. Iron overload frequently occurs in individuals with thalassemia or other transfusion-dependent conditions, as each transfusion unit contributes an additional 200–250 mg of iron to the body (3, 4). Notably, the liver serves as a central hub for iron metabolism and homeostasis.
Among various organs, the liver plays a key role in regulating iron metabolism and maintaining systemic iron balance (5). Its primary functions include storing excess iron and controlling the amount of iron released into circulation (6). This process is mainly governed by the production and release of hepcidin, a peptide hormone that modulates iron entry into the bloodstream (7). Hepcidin stimulates the degradation of ferroportin, an iron exporter, thereby regulating iron release from cells (8). Furthermore, the liver participates in the absorption of dietary iron via Divalent Metal Transporter 1 (Dmt1) and Transferrin Receptor 1 (Tfr1). A key function of Tfr1 in maintaining iron homeostasis is its interaction with the hemochromatosis protein Hfe, which is critical for regulating hepcidin production (9).
Several hepatic genes are critical in iron metabolism, including Tfr1, Hepcidin Antimicrobial Peptide (Hamp), and Ferritin Light Chain (Ftl). The Hamp gene encodes hepcidin, the principal hormone governing iron homeostasis. Hepcidin expression increases in response to elevated iron levels or inflammatory stimuli, which is commonly associated with hyperferritinemia (10). Conversely, in healthy individuals, hepcidin production decreases in response to low iron levels and other stimuli (8). Ferritin, encoded in part by Ftl, is an intracellular iron-storage protein, and its serum levels are widely used as a surrogate marker for body iron status, increasing during iron overload (11). Serum ferritin levels correlate directly with total body iron stores, making it a valuable biomarker for estimating iron status (12, 13). Meanwhile, Tfr1 facilitates cellular iron uptake and can become saturated in individuals with HFE gene mutations (14). To prevent iron accumulation and its detrimental effects, iron chelation therapy is employed as an effective intervention.
Iron chelation therapy is the primary treatment for iron overload, utilizing pharmacological agents such as Deferiprone (DFP), Deferoxamine (DFO), and Deferasirox (DFX) to bind excess circulating iron (15). Beyond reducing iron burden, these agents also suppress ROS formation, thereby mitigating oxidative damage (16). Among available chelators, DFP is particularly effective at reducing intracellular labile iron and curbing oxidative stress (17). Its mechanism involves chelating Fe²⁺ ions at the active site of iron-dependent histone lysine demethylases, inhibiting their enzymatic function (18).
This study aims to evaluate the expression of Hamp, Ftl, and Tfr1 genes in the liver of rats under normal and iron overload conditions. Additionally, it investigates the effects of DFP administration on the expression levels of these genes in a rat model of iron overload.
Experimental Section
Materials
The materials used in this study included Deferiprone, Iron Dextran (Hemadex; Sanbe Farma, Bandung, Indonesia), RNA isolation kit (Quick-RNA™ Miniprep Plus Kit, Zymo Research), cDNA synthesis kit (FIREScript RT cDNA Synthesis Kit, Solis BioDyne), and primers (Macrogen).
In Vivo Test
Animal Preparation
This study was approved by the Animal Research Ethics Committee of Universitas Padjadjaran, Indonesia (Approval No. 75/UN6. KEP/EC/2023). A total of 18 healthy male Wistar rats (150–200 g) were housed in standard cages under controlled environmental conditions, with a 12-hour light/dark cycle. Food and water were provided ad libitum throughout the study (19). Animals were acclimatized for 7 days prior to experimental procedures.
Sample Administration and Liver Collection
Rats were randomly divided into three groups (n = 6 per group): Normal (N), Negative Control (NC, Iron Dextran), and Treatment (T, Iron Dextran + DFP). The sample size was calculated using Mead’s Resource Equation method (E = N − B − T) to ensure both ethical animal use and sufficient statistical power. Iron dextran (120 mg/kg BW) was administered intravenously every 3 days for a total of 15 days (20). The Treatment group received DFP orally at a dose of 100 mg/kg BW/day, divided into three equal doses, for 28 consecutive days. At the end of the treatment period, rats were fasted for 16 h and anesthetized via intramuscular injection of Ketamine-Xylazine (0.2 mL). Liver tissues were collected, rinsed with physiological saline, weighed, and stored at −20°C for subsequent molecular analysis.
RNA Isolation and Extraction
Total RNA was extracted from liver tissues using the Quick-RNA™ Miniprep Plus Kit (Zymo Research), following the manufacturer’s instructions. The protocol included lysis buffer preparation, tissue homogenization, and RNA purification. RNA integrity and purity were assessed using a Nanodrop spectrophotometer, with acceptable purity defined by a 260/280 absorbance ratio of 1.8–2.0.
cDNA Synthesis and RT-PCR
First-strand cDNA synthesis was performed using the FIREScript RT cDNA Synthesis Kit (Solis BioDyne) according to the manufacturer’s protocol. RT-PCR conditions were optimized by testing annealing temperatures between 55–65°C. Amplification was carried out using the GENECHECKER UF-300 real-time thermal cycler. The thermal cycling protocol is detailed in Table 1. Primer sequences were synthesized by Macrogen, and the complete list of primer pairs used in the analysis is presented in Table 2.
| Step | Temperature (oC) | Time | Cycles |
|---|---|---|---|
| Initial Activation | 95 | 12 min | 1 |
| Denaturation | 95 | 15 s | |
| Annealing | 58 | 20 s | 40 |
| Extension | 72 | 20 s |
| Gene | Primer Sequence 5’ 3’ | Tm (oC) | Ref. |
|---|---|---|---|
| Hamp | Forward: 5’-CTG CCT GTC TCC TGC TTC TCC-3’ | 65.9 | (21) |
| Reverse: 5’-AGT TGG TGT CTC GCT TCC TTC G-3’ | 64.2 | ||
| Ftl | Forward: 5’-GCT GGC TTC TTG ATG TCC-3’ | 58.5 | |
| Reverse: 5’-CCT CCT ACA CCT ACC TCT C-3’ | 59.7 | ||
| Tfr1 | Forward: 5’-AGT CAT CTG GAT TGC CTT CTA TAC C-3’ | 60.1 | |
| Reverse: 5’-TAG CGG TCT GGT TCC TCA TAG C-3’ | 63.1 | ||
| Gapdh | Forward: 5′-CCATCAACGACCCCTTCATT-3′ | 58 | (22) |
| Reverse: 5′-CACGACATACTCAGCACCAGC-3′ | 58 |
Data Analysis
Cycle threshold (Ct) values obtained from RT-PCR for the Normal (N), Negative Control (NC), and Treatment (T) groups were analyzed. Relative expression (RE) was calculated using the Livak method: RE = 2^−ΔΔCt (14). Statistical analyses were performed using GraphPad Prism 9 software. Data were first assessed for normality using the Shapiro–Wilk test. For normally distributed data, a one-way analysis of variance (ANOVA) followed by Dunn’s multiple comparisons test was used to evaluate significant differences. All analyses were conducted at a 95% confidence level (p < 0.05), and results are presented graphically.
Results and Discussion
Relative Expression of mRNA Hamp
Quantitative RT-PCR analysis demonstrated that administration of Iron Dextran (ID) significantly elevated hepatic Hamp mRNA expression. Cycle threshold (Ct) values obtained from liver tissue RT-PCR were used to assess gene expression across the three experimental groups. Relative expression (RE) was calculated using the 2^−ΔΔCt method described by Livak and Schmittgen (2001), with results illustrated in Figure 1. This approach allows normalized comparison of gene expression among the experimental groups.
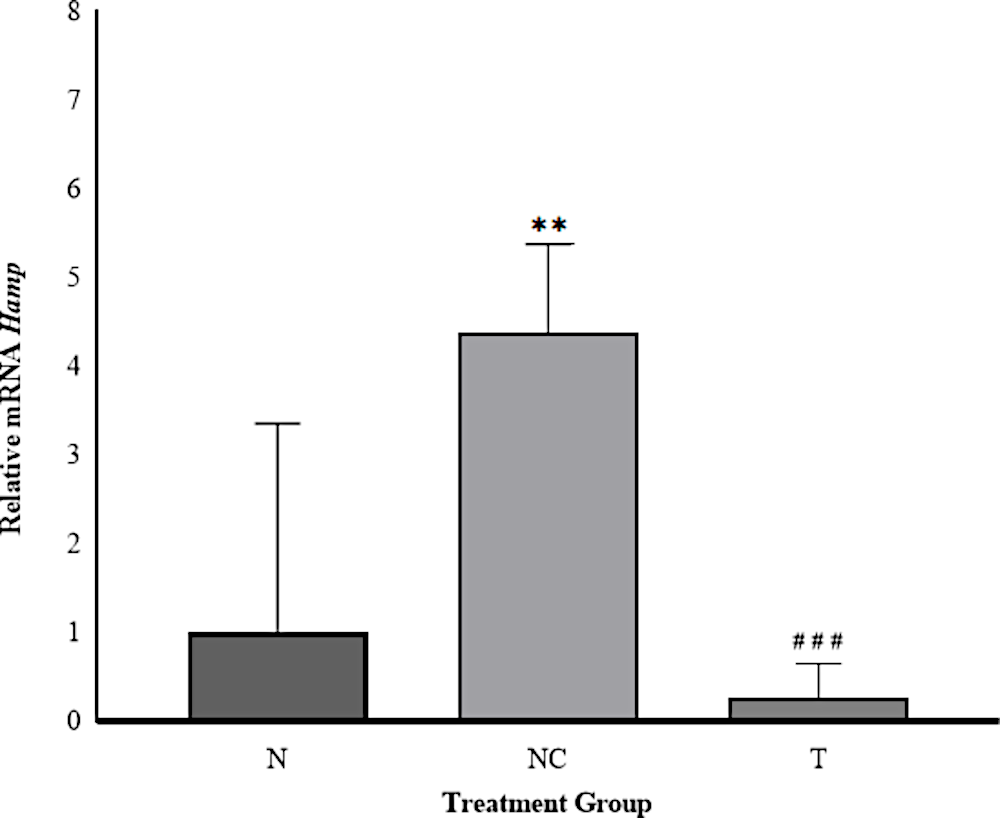
In the NC group, Hamp expression increased 4.39-fold relative to the N group, indicating substantial upregulation in response to iron overload. This aligns with the physiological upregulation of hepcidin under conditions of excessive hepatic iron deposition (21, 22). Hepcidin synthesis is modulated by multiple regulatory signals, including systemic iron levels, inflammation, and erythropoietic activity, with iron predominantly acting via the Bone Morphogenetic Protein (BMP)/Smad pathway. In this mechanism, BMPs bind to BMP receptors (BMPRs) and form complexes with the co-receptor hemojuvelin (HJV), triggering Smad phosphorylation and transcriptional activation of Hamp (23, 24).
Rats in the T group exhibited a marked reduction in Hamp expression (~0.27-fold relative to the N group), representing a 93.85% decrease compared to the NC group. This suggests that DFP administration effectively attenuated hepcidin gene transcription, consistent with previous reports demonstrating DFP’s capacity to reduce hepatic iron burden and suppress Hamp expression (25). Under controlled iron levels, hepatic Hamp expression remains low, permitting normal iron absorption and systemic distribution (21). This pathway-mediated induction of hepcidin represents a key adaptive response to iron overload aimed at maintaining systemic iron balance.
Relative Expression of mRNA Ftl
Ftl gene expression among experimental groups is shown in Figure 2. The N group served as the control, normalized to a relative expression value of 1.00.
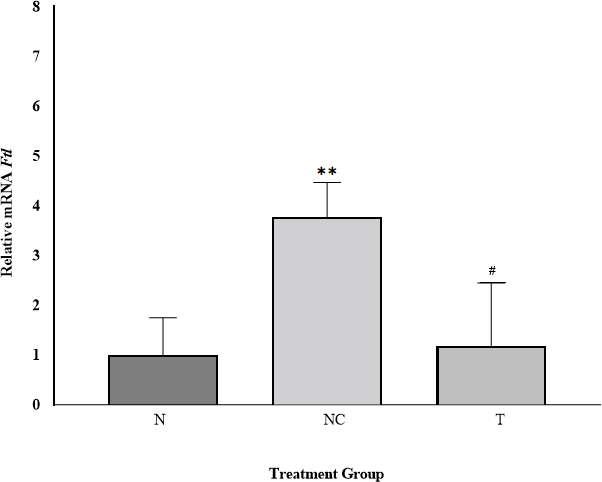
Ftl expression in the NC group increased approximately 3.77-fold relative to N, consistent with hepatic iron accumulation (26, 27, 28). In the T group, Ftl expression was only modestly elevated (~1.18-fold relative to N), indicating normalization following DFP treatment. Compared with the NC group, Ftl expression in the T group decreased by 0.31-fold, demonstrating effective downregulation toward baseline levels. This mirrors clinical observations where DFP monotherapy significantly reduces serum ferritin levels in transfusion-dependent thalassemia patients (29, 30, 31).
Relative Expression of mRNA Tfr1
Tfr1 expression across the experimental groups is illustrated in Figure 3. Expression values were normalized to the N group (set at 1.00) to enable fold-change comparisons.
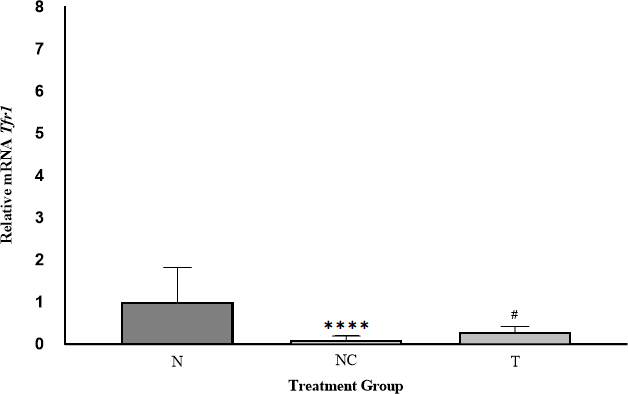
In the NC group, Tfr1 expression was 0.09-fold relative to N, reflecting diminished interaction between Iron Regulatory Proteins (IRPs) and Iron Responsive Elements (IREs) under iron overload (32, 33). This post-transcriptional regulation via the IRE-IRP system inversely correlates with cellular iron status. Following DFP administration, Tfr1 expression increased (~0.29-fold relative to N), indicating normalization of gene expression. The relative Ftl expression in the T group increased 3.22-fold compared with the NC group, highlighting DFP’s role in reducing iron-mediated toxicity by removing iron from transferrin and non-transferrin-bound iron (NTBI) pools (34, 35, 36).
Overall, DFP treatment effectively reversed iron-induced dysregulation of Hamp, Ftl, and Tfr1 expression, demonstrating its efficacy in modulating hepatic iron-related gene expression under iron overload conditions (Table 3). These coordinated transcriptional changes further support the role of DFP in restoring hepatic iron homeostasis through normalization of iron sensing and transport mechanisms.
| Gene | NC vs Normal | T vs NC |
|---|---|---|
| Hamp | ↑ Upregulated | ↓ Downregulated |
| Ftl | ↑ Upregulated | ↓ Downregulated |
| Tfr1 | ↓ Downregulated | ↑ Upregulated |
Conclusion
This study demonstrates that iron overload induces significant dysregulation of hepatic iron-regulatory genes, characterized by upregulation of Hamp and Ftl and downregulation of Tfr1. Treatment with Deferiprone (DFP) effectively reversed these alterations by suppressing Hamp and Ftl expression while restoring Tfr1 levels toward those observed in the normal group. These findings indicate that DFP promotes the re-establishment of hepatic iron homeostasis at the molecular level and may be beneficial for managing iron overload–associated liver dysfunction.
Declarations
Acknowledgment
The author respectfully acknowledge Prof. Dr. Ratu Safitri, MS. for generous financial support and providing throughout the course of this research. The author are also deeply grateful to Dr. Yasmi Purnamasari Kuntana, MP. and Mrs. Tanendri Arrizqiyani, M.Si., for their critical guidance, insightful feedback and continuous supervision, all of which greatly contributed to the successful completion of this study
Conflict of Interest
The authors declare no conflicting interest.
Data Availability
The unpublished data is available upon request to the corresponding author.
Ethics Statement
This research received ethical approval from the Animal Research Ethics Committee of Universitas Padjadjaran, Bandung, under approval number 75/UN6.KEP/EC/2023. All procedures involving experimental animals were conducted in strict accordance with the ethical guidelines established by Universitas Padjadjaran, based on internationally accepted principles for the humane treatment of animals.
Funding Information
The author(s) declare that no financial support was received for the research, authorship, or publication of this article.
References
- Zaychenko GV, Gorchakova N, Shumeiko OV, Klymenko OV. Iron: biochemical, pharmacological, and clinical data. Bull Probl Biol Med. 2022;3(166):36.
- Fianza PI, Rahmawati A, Widihastha SH, et al. Iron overload in transfusion-dependent Indonesian thalassemic patients. Anemia. 2021;1(2021):1–9.
- Hsu CC, Senussi NH, Fertrin KY, Kowdley KV. Iron overload disorders. Hepatol Commun. 2022;6(8):1842–54.
- Scannell C. In transfusion-dependent thalassemia children, increased iron overload is associated with lower serum alpha-Klotho, which is strongly associated with lower total and ionized calcium concentrations. Curr Mol Med. 2023;23(5):442–52.
- Li Y, Huang X, Wang J, et al. Regulation of iron homeostasis and related diseases. Hindawi. 2020;1(2020)1-11.
- Ameka M, Hasty AH. Paying the iron price: liver iron homeostasis and metabolic disease. Compr Physiol. 2022;12(3):3641–63.
- Bloomer SA, Brown KE. Hepcidin and iron metabolism in experimental liver injury. Am J Pathol. 2021;199(5):1165–79.
- Wang C, Babitt JL. Liver iron sensing and body iron homeostasis. Blood. 2019;122(1):18–29.
- Xiao X, Moschetta GA, Xu Y, et al. Regulation of iron homeostasis by hepatocyte TFR1 requires HFE and contributes to hepcidin suppression in β-thalassemia. Blood. 2022;141(4):422–32.
- Faraj SA, Al-Abedy NM. Hepcidin. In: IntechOpen; 2022.
- Nurbahiyah E, Maulina D. Profil penggunaan obat kelasi besi pada pasien talasemia poli anak di Rumah Sakit X Jatinegara. Indones J Health Sci. 2023;3(2):149-154.
- Arifie R, Syamsunarno MRAA, Panigoro R. Serum ferritin sebagai parameter status besi tubuh. Indones J Clin Pathol Med Lab. 2022;28(1):57–61.
- Vázquez LI, Arija V, Aranda N, et al. Factors associated with serum ferritin levels and iron excess: results from the EPIC-EurGast study. Eur J Nutr. 2021;60(1):101-114.
- Kim H. Diagnosis and treatment of transfusion-related iron overload. J Korean Med Assoc. 2022;65(10):662–72.
- Bruzzese A, Martino EA, Mendicino F, et al. Iron chelation therapy. Eur J Haematol. 2023;110(5):490–7.
- Carter A, Racey S, Veuger S. The role of iron in DNA and genomic instability in cancer: a target for iron chelators that can induce ROS. Appl Sci. 2022;12(19):10161.
- Zheng R, Fan Y, Gadde R, Betharia S, Kim J. Deferiprone decreases labile iron in brain and ameliorates abnormal emotional behavior in mice with brain iron accumulation. Neuropharmacology. 2023;385:340.
- Khodaverian V, Tapadar S, MacDonald IA, et al. Deferiprone: pan-selective histone lysine demethylase inhibition activity and structure–activity relationship study. Sci Rep. 2019;9(1):4802.
- Badan Pengawas Obat dan Makanan Republik Indonesia. Peraturan Badan Pengawas Obat dan Makanan Nomor 18 Tahun 2021 tentang Pedoman Uji Farmakodinamik Praklinik Obat Tradisional. Jakarta: BPOM; 2021.
- Zhang H, Pavel Z, Shaohua W, Gavin YO. Role of iron metabolism in heart failure: from iron deficiency to iron overload. Biochim Biophys Acta Mol Basis Dis. 2019;1865(7):1925–37.
- Kowdley DS, Kowdley KV. Appropriate clinical genetic testing of hemochromatosis type 2–4, including ferroportin disease. Appl Clin Genet. 2021;14:353-361.
- Prasnicka A, Lastuvkova H, Cermanova A, et al. Iron overload reduces synthesis and elimination of bile acids in rat liver. Sci Rep. 2019;9(1):9780.
- Liu J, Sun B, Yin H, Liu S. Hepcidin: a promising therapeutic target for iron disorders. Medicine (Baltimore). 2016;95(14):e3150.
- Silvestri L, Pagani A, Nai A, et al. The serine protease matriptase-2 (TMPRSS6) inhibits hepcidin activation by cleaving membrane hemojuvelin. Cell Metab. 2008;8(6):502–11.
- Kontoghiorghes GJ. The vital role played by deferiprone in the transition of thalassaemia from a fatal to a chronic disease and challenges in its repurposing for use in non-iron-loaded diseases. Pharmaceuticals. 2023;16(7):1016.
- Vulinović MP, Turčić P, Micek V, Ljubojević M. Light and heavy ferritin chain expression in the liver and kidneys of Wistar rats: aging, sex differences, and impact of gonadectomy. Arch Ind Hyg Toxicol. 2022;73(1):48–61.
- Panigoro R, Ahmad FM, Gamayani U, Anggraeni N, Widyastuti R, Ghozali M, Syamsunarno MRAA. Acute iron dextran injection increases liver weight and reduces glycerol kinase expression in liver. Indones J Clin Pharm. 2018;7(4):236-242.
- Anderson GJ, Frazer DM. Current understanding of iron homeostasis. Am J Clin Nutr. 2017;106(Suppl 6):1559S-1566S.
- Jahir SJ, Anwar S, Morshed AA, et al. Comparative study of deferiprone versus deferasirox as iron chelation therapy in thalassemic children. Bangladesh J Child Health. 2020;44(3):152–6.
- Kittipoom T, Tantiworawit A, Punnachet T, et al. The long-term efficacy of deferiprone in thalassemia patients with iron overload: real-world data from the registry database. Hemoglobin. 2022;46(2):75–80.
- Binding A, Ward R, Tomlinson G, Kuo KHM. Deferiprone exerts a dose-dependent reduction of liver iron in adults with iron overload. Eur J Haematol. 2019;103(2):80–87.
- Wessling-Resnick M. Crossing the iron gate: why and how transferrin receptors mediate viral entry. Annu Rev Nutr. 2018;38:431–458.
- Recalcati S, Gamella E, Buratti P, Cairo G. Molecular regulation of cellular iron balance. IUBMB Life. 2017;69(6):389–98.
- Kontoghiorghe CN, Kolnagou A, Kontoghiorghes GJ. Potential clinical applications of chelating drugs in diseases targeting transferrin-bound iron and other metals. Expert Opin Investig Drugs. 2013;22(5):591–618.
- Miyazawa M, Bogdan AR, Hashimoto K, Tsuji Y. Regulation of transferrin receptor-1 mRNA by interaction between iron-responsive element-binding proteins and miR-7/miR-141 in the 3′-IRE stem-loop. RNA. 2018;24(4):468–79.
- Kontoghiorghes GJ, Kleanthous M, Kontoghiorghe CN. The history of deferiprone (L1) and the paradigm of the complete treatment of iron overload in thalassemia. Mediterr J Hematol Infect Dis. 2020;12(1):e2020011.