Mangrove ecosystems, distributed across tropical and subtropical coastlines, are globally recognized for their exceptional biodiversity and multifunctional ecological roles. They sustain coastal fisheries by serving as breeding and nursery habitats, support diverse terrestrial and aquatic fauna, enhance shoreline stability through wave attenuation and erosion control, and improve water quality via nutrient recycling and pollutant filtration. Mangroves are also among the most efficient blue carbon sinks, contributing significantly to climate-change mitigation. From a socio-economic perspective, they provide timber and non-timber resources, support artisanal fisheries, enable ecotourism-based livelihoods, and hold high potential for carbon-credit revenue and bioprospecting. The review synthesizes evidence from 135 studies published between 1990 and 2024, retrieved from Web of Science, Scopus, Google Scholar, and institutional repositories, based on their relevance to mangrove ecology, ecosystem services, and community-level economic benefits. Only peer-reviewed articles, technical reports, and case studies with clear methodological descriptions were included. The synthesis reveals that the ecological and economic functions of mangroves are strongly interdependent, with degradation of habitat quality directly reducing fisheries productivity, carbon storage efficiency, and livelihood security. The review highlights the urgent need for integrated management approaches, including hydrological restoration, community-based conservation, and policy-supported payment-for-ecosystem-services mechanisms, to enhance ecosystem resilience under accelerating anthropogenic and climate pressures.
Mangroves are among the world’s most productive and ecologically significant ecosystems. These salt-tolerant trees and shrubs occupy intertidal zones of tropical and subtropical coastlines, where they provide a wide array of ecological and economic services despite covering only about 0.1% of Earth’s land area (1, 2). Their structurally complex habitats support exceptionally high biodiversity and function as critical interfaces between terrestrial and marine environments. By linking these systems, mangrove forests facilitate species movement, nutrient exchange, and energy flow, thereby contributing substantially to coastal ecological integrity and human well-being (3, 4).
Ecologically, mangroves regulate climate by acting as nursery habitats for aquatic fauna, stabilizing coastlines, buffering storm surges, and storing large quantities of atmospheric carbon (5, 6). Economically, they provide cost-effective natural protection from coastal hazards while supporting fisheries, timber and non-timber forest products, and a variety of nature-based livelihoods. Despite these benefits, mangroves are under increasing pressure from multiple anthropogenic and climatic drivers. However, these threats vary significantly across regions: shrimp aquaculture and land conversion dominate losses in Southeast Asia, coastal infrastructure expansion is a primary driver in South Asia and West Africa, while accelerated subsidence and sea-level rise disproportionately affect small island nations and deltaic regions (9, 3). Understanding these spatial and thematic variations is essential for effective conservation.
Although mangrove ecosystems have been widely studied, current research reveals important knowledge gaps. Existing literature is often regionally biased, with heavy concentration in Southeast Asia, creating limited global comparability. Considerable uncertainty persists regarding key ecological processes such as the magnitude of detrital outwelling, regional differences in carbon stock estimates, and variation in restoration success under different hydrological conditions. Similarly, economic assessments lack standard valuation frameworks, limiting their integration into policy instruments such as carbon markets, coastal planning, and payment-for-ecosystem-services programs.
Given these gaps, this review aims to provide a comprehensive, integrative synthesis of the ecological functions and socio-economic significance of mangroves at a global scale. Specifically, it aims to: 1) Compare ecological processes across different biogeographic regions, highlighting convergences and divergences. 2) Critically evaluate contrasting evidence-particularly in areas such as carbon dynamics, outwelling, and hydrological resilience. 3) Synthesize economic valuations of mangrove services, including fisheries, NTFPs, ecotourism, carbon credits, and emerging biotechnological applications. 4) Identify research gaps and policy challenges relevant to conservation, restoration, and climate adaptation.
By addressing these objectives, this review moves beyond descriptive accounts to provide a structured, evidence-based evaluation of mangrove ecosystem services, emphasizing their importance for long-term environmental stability, coastal resilience, and sustainable community livelihoods.
Methodology
A comprehensive literature search was conducted between January and July 2024 across Web of Science, Scopus, ScienceDirect, Google Scholar, SpringerLink, and institutional repositories such as FAO, UNEP and IUCN, using combinations of keywords including “mangroves,” “ecological functions,” “blue carbon,” “nursery habitats,” “coastal protection,” “economic valuation,” “NTFPs,” “ecotourism,” and “aquaculture impacts. ” The search covered studies published between 1990 and 2024. Articles were included if they examined ecological or economic aspects of mangrove ecosystems, presented original data or validated analyses, described clear methodology, and were available in English, whereas anecdotal reports, non-reviewed sources, conference abstracts without full text, and studies focused solely on unrelated wetland types were excluded. A total of 312 records were identified, 72 duplicates were removed, and 230 full texts were screened; ultimately, 135 studies fulfilled all inclusion criteria. Extracted data were synthesized thematically and classified into ecological and economic domains. Blue Carbon frameworks to ensure structural consistency. The methodology also explicitly acknowledges publication bias (positive results being overrepresented), regional citation bias heavily favouring Southeast Asia, and methodological differences across studies (e. g. , field plots vs. remote sensing) that influence comparability. A PRISMA-style flow diagram documenting identification, screening, eligibility assessment, and inclusion steps was prepared for transparency (10), as shown in Figure 1.
Figure 1. PRISMA flow diagram outlining the identification, screening, eligibility assessment, and inclusion of studies in this review.
Ecological Significance
Mangrove ecosystems provide a wide array of essential ecological services that underpin biodiversity, protect coastlines, regulate water quality, and store significant quantities of carbon. Their unique structural adaptations such as aerial roots, pneumatophores, prop roots, dense canopies, and high sediment-trapping capacity create complex, layered habitats that support diverse terrestrial, estuarine, and marine communities. As biodiversity hotspots, mangroves offer critical breeding, feeding, and nursery grounds for fish, crustaceans, birds, and other wildlife.
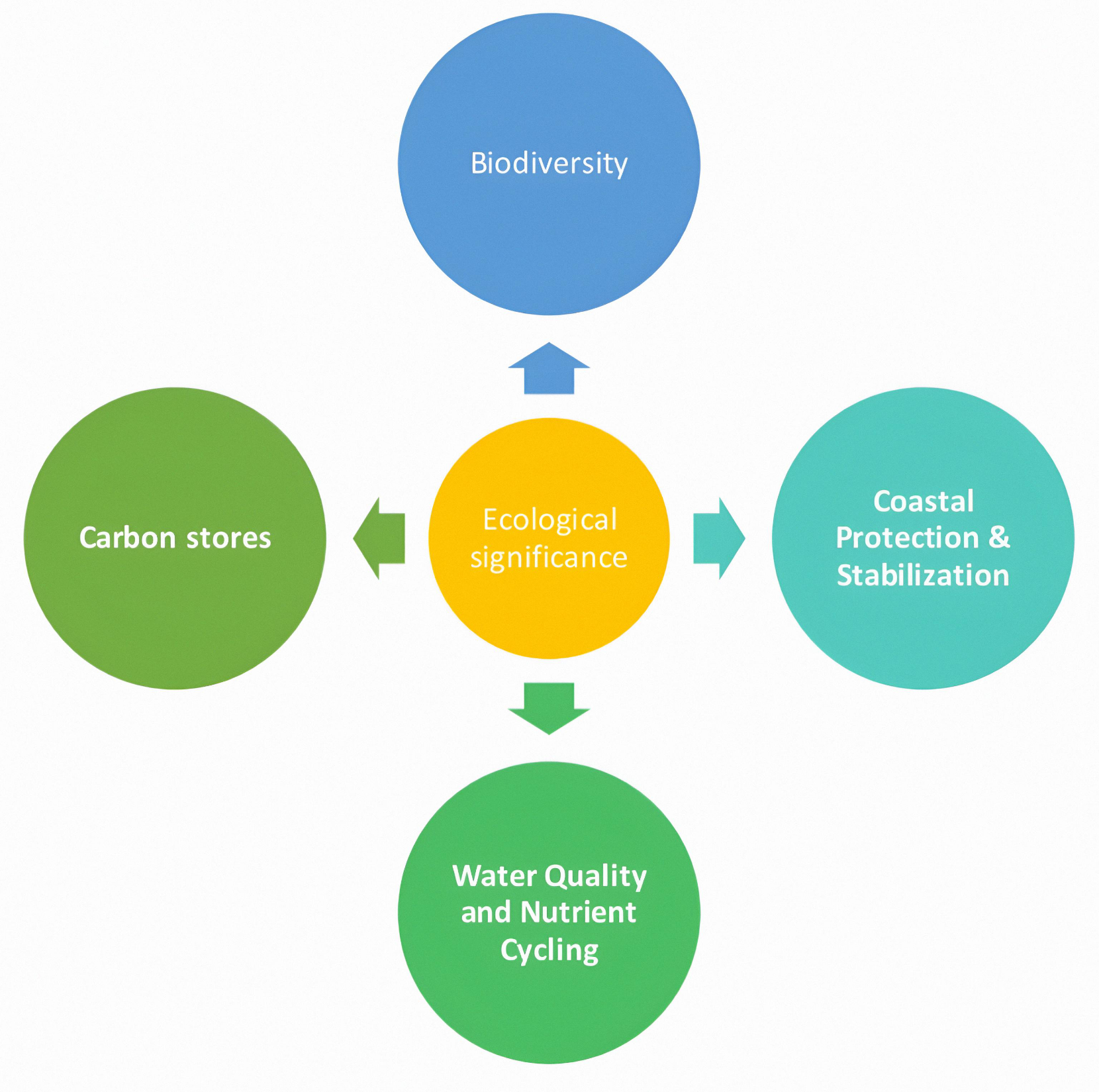
Globally, mangroves are regarded as some of the most ecologically valuable habitats, functioning as natural buffers that absorb wave energy, reduce storm impacts, and prevent shoreline erosion. They also enhance water quality by trapping sediments, immobilizing heavy metals, and filtering pollutants before these reach open waters. Importantly, mangroves serve as highly efficient carbon sinks, storing large amounts of organic carbon in both biomass and sediments, thereby contributing significantly to global climate regulation, as shown in Figures 2 and 3.
Figure 2. Diagram showing ecological significance of Mangroves.
Figure 3. Different ecological role of Mangroves.
Biodiversity Conservation
Mangrove forests function as vital biodiversity hotspots in tropical and subtropical coastal regions, supporting remarkable terrestrial and aquatic species diversity through their unique structural complexity. Features such as dense prop roots, pneumatophores, and stratified canopies create multiple microhabitats that sustain rich and interconnected ecological communities (3, 4, 11). One of the most critical ecological functions of mangroves is their role as breeding, nursery, and rearing grounds for a wide range of marine organisms of both ecological and economic importance (3, 4, 11). Juvenile stages of numerous fishes, shrimps, crabs, and mollusks depend on the protective structure of mangrove roots for shelter, predator avoidance, and foraging, significantly increasing their chances of survival (12, 13). Empirical studies indicate that nearly 75% of commercially harvested tropical fish species utilize mangrove habitats during some phase of their life cycle, highlighting their indispensable contribution to coastal fisheries productivity (14, 15).
The structural complexity of mangrove habitats also supports high shrimp productivity (16), including species of major commercial value such as tiger prawns (Penaeus monodon) and sea crabs (Scylla paramamosain), as well as brackish-water fishes like barramundi (Lates calcarifer) and Nile tilapia (Oreochromis niloticus) (17).
Mangroves provide essential habitats for a wide array of resident and migratory bird species including herons, egrets, kingfishers, storks, sea eagles, and ospreys serving as nesting, roosting, and stopover sites along major coastal flyways (18-20). Across biogeographical regions, between 150 and 250 bird species have been documented within mangrove landscapes (21).
Beyond avifauna, mangroves sustain a diverse assemblage of mammals (e. g. , proboscis monkeys Nasalis larvatus, fishing cats Prionailurus viverrinus), reptiles (including crocodiles and snakes), and invertebrates such as fiddler crabs and mangrove tree crabs that play crucial roles in nutrient cycling (22, 23). Southeast Asia, recognized as the global epicentre of mangrove biodiversity, exhibits particularly high species richness and endemism (4). For example, Malaysia’s approximately 9, 000 hectares of mangrove forests support extensive marine and estuarine life, including seagrasses (Gracilaria spp. ), algae, fungi, and diverse fish communities, with species distributions influenced by tidal dynamics and seasonal salinity patterns (24). These ecosystems also enhance ecotourism potential, notably illustrated by the famous firefly (Photuris lucicrescens) congregations along the Kampung Kuantan riverbanks in Malaysia (25). Toward temperate intertidal zones, mangroves continue to support burrowing species such as clams, mussels, fiddler crabs, and polychaetes (17).
Coastal Protection & Stabilization
Mangroves function as highly effective natural coastal defence systems through a combination of hydrodynamic and geomorphological mechanisms. Their dense vegetation and complex root architectures exhibit strong wave-attenuation capacity, reducing wave energy by 13–90% over a 100-meter mangrove belt (26-28) and lowering storm-surge heights by approximately 5–50 cm per kilometre of forest width (29). These protective benefits were clearly demonstrated during Hurricane Wilma (2005), when mangroves in Florida reduced inland flooding by nearly 70% (30), and following the 2004 Indian Ocean tsunami, during which mangrove belts absorbed up to 70% of incoming wave energy (31, 32).
Beyond wave reduction, mangrove root systems significantly enhance shoreline stability by trapping sediments and facilitating vertical accretion. They can retain up to 80% of incoming sediments (33) and contribute to long-term elevation gain through sediment accumulation processes (34, 35). These functions are particularly critical in erosion-prone zones such as Vietnam and Malaysia, where coastal erosion can reach 50 m/year and currently affects nearly 30% of shoreline segments (28, 36).
In addition to physical protection, mangroves provide economically viable alternatives to engineered coastal barriers. For instance, Philippine mangroves offer an estimated USD 1.6 billion in annual storm-damage prevention (6, 36). Their proven cost-effectiveness, coupled with demonstrated performance during extreme climatic events (37), underscores the value of mangrove conservation as a nature-based solution for enhancing coastal resilience an increasingly urgent need as climate change drives higher storm frequency and intensity (38).
Water Quality and Nutrient Cycling
Mangrove ecosystems play a crucial role in maintaining coastal water quality through their ability to trap pollutants, retain excess nutrients such as nitrogen and phosphorus, and capture suspended solids from agricultural and urban runoff. Their intricate root structures and sediment-binding capacity significantly reduce turbidity and prevent eutrophication in adjacent ecosystems, thereby protecting sensitive habitats such as seagrass beds and coral reefs from sedimentation and nutrient overload (39, 8). By stabilizing sediments and minimizing water cloudiness, mangroves enhance seagrass growth parameters including shoot density, root length, and overall biomass while supporting the health of nearby coral reef systems.
Mangroves also contribute substantially to regional biogeochemical cycles and function as integral nodes in the tropical coastal blue carbon network. These ecosystems store large amounts of organic carbon in both biomass and sediments, reinforcing the need for integrated conservation approaches across interconnected mangrove–seagrass–coral reef systems (41, 42, 44, 45).
The long-standing hypothesis that mangrove-derived detritus supports offshore productivity has been revisited in recent years. While traditional models emphasize strong detrital “outwelling,” new mass-balance and tracer studies suggest that export may be less extensive than previously assumed. Dissolved organic matter appears to play a more significant role than particulate detritus in fueling adjacent food webs, and benthic communities show variable dependence on mangrove-derived detrital inputs (46).
Microbial processes within mangrove soils further enhance nutrient cycling. Microbial communities facilitate nitrogen fixation, denitrification, and organic matter decomposition, thereby increasing nutrient bioavailability, supporting plant growth, and suppressing pathogenic bacteria ultimately promoting higher biodiversity across the ecosystem (47). Through these mechanisms, mangroves function as effective nutrient processors, recycling excess terrestrial inputs and preventing nutrient accumulation that could otherwise trigger harmful algal blooms (48). Detritus from leaf litter and organic debris forms the foundation of complex food webs within and around mangrove ecosystems (49).
Importantly, mangrove sediments act as sinks for heavy metals and organic contaminants through adsorption and plant uptake, providing a natural buffer against anthropogenic pollution (50). This function is especially vital in regions exposed to intense agricultural runoff, aquaculture effluents, and urban wastewater discharge, where mangroves mitigate ecological degradation and help maintain coastal water quality.
Carbon Stores
Mangrove forests span 105 countries, with Indonesia, Brazil, Malaysia, and Papua New Guinea together holding nearly half of the global mangrove area. These ecosystems are among the most carbon-dense in the tropics, storing substantially larger quantities of carbon per unit area than most terrestrial forests, making them indispensable for climate-change mitigation (51). Studies across Southeast Asia particularly from Indonesia consistently report some of the highest carbon stocks worldwide, with values commonly ranging between 863–1073 Mg C ha⁻¹ in mature stands supported by large trees and deep organic sediments (52).
A broader synthesis reveals a coherent pattern: structurally complex, undisturbed mangroves function as powerful long-term carbon sinks, with annual sequestration rates often exceeding those of many tropical terrestrial forests and capturing more than 2 tons of carbon per hectare per year (53, 66). Quantifying these pools remains critical for evaluating mangrove contributions to global ecosystem services (7). More than 80% of this carbon typically resides below ground, stored in sediments and dense root systems (21, 7, 54). Soil carbon accumulation driven by litterfall, woody debris, and slow decomposition is strongly modulated by forest structure (tree height, basal area) and hydrological conditions such as salinity and dissolved oxygen (54, 7).
Although intact mangroves act as efficient, long-term carbon sinks, their degradation rapidly reverses this role. Disturbance exposes deep anoxic sediments to oxygen, accelerating decomposition and releasing substantial CO₂; for instance, conversion of mangroves to pasture generates emissions far higher than equivalent deforestation in the Amazon (55). Nevertheless, remaining intact mangroves still offset the majority of emissions from global mangrove loss up to 94% (55).
Global assessments estimate that mangroves collectively store approximately 21, 896.56 Mt CO₂-equivalent, with 2, 817.23 Mt contained in above-ground biomass and 19, 079.32 Mt within the top 1 m of soil (59, 60). National inventories indicate that Indonesia holds the highest stock (61). Soil carbon typically extends 0.5–3 m deep, accounting for 49–98% of total ecosystem carbon (62).
Importantly, differences in reported values across studies often reflect contrasting methodological approaches. Local case studies such as plot-based measurements from Indonesia use direct biomass sampling and sediment coring, providing fine-scale, high-resolution estimates but limited spatial coverage. In contrast, global carbon stock assessments rely heavily on remote sensing, upscaled biomass models, and harmonised datasets, which capture large-scale patterns but may smooth out local heterogeneity. Recognising these methodological differences helps reconcile apparent variation in reported values and underscores the need for integrative approaches combining field data with remote-sensing products to improve regional and global carbon accounting.
Economic Significance
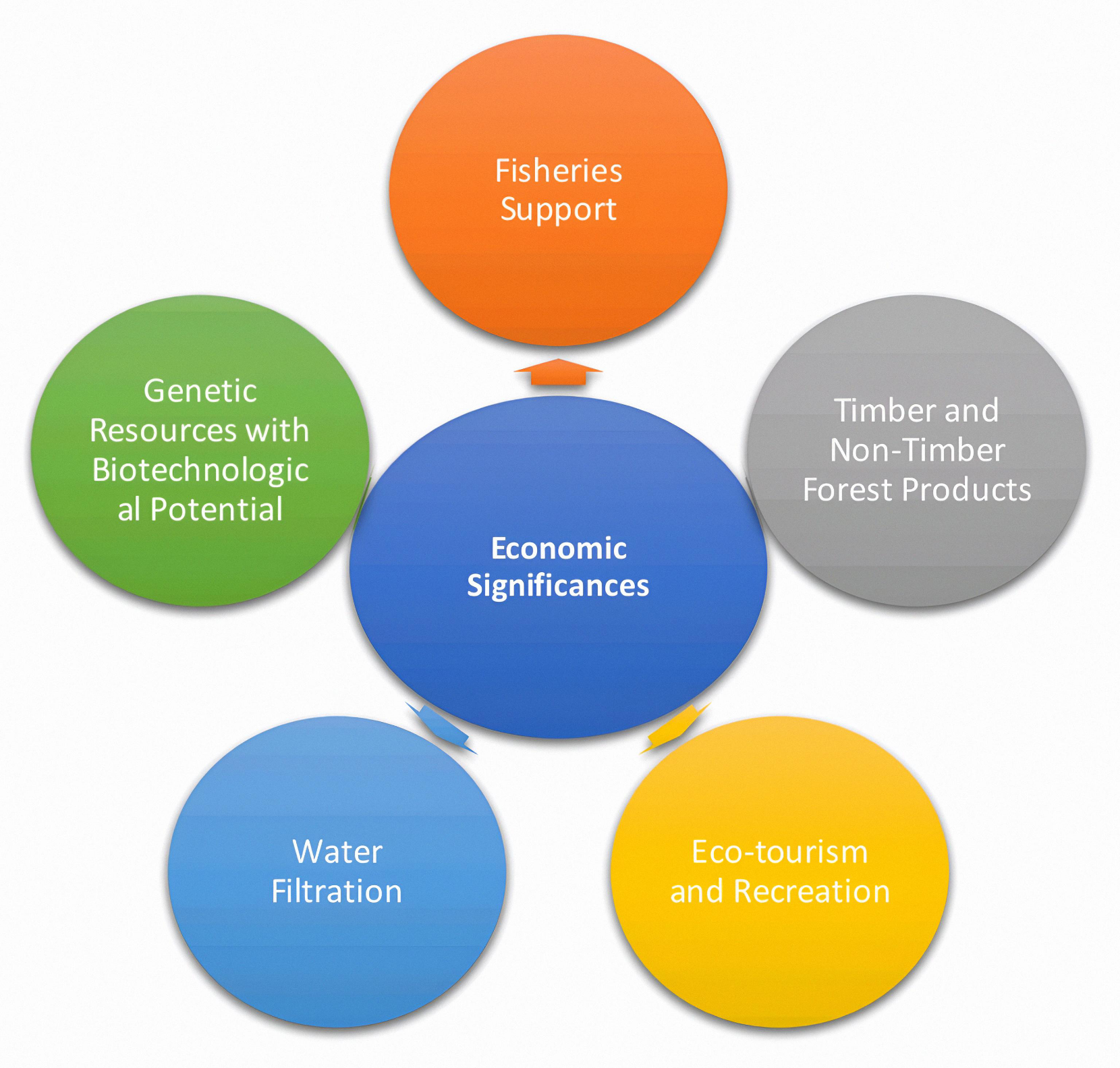
Mangroves hold immense economic importance by supporting fisheries, aquaculture, timber, fuelwood, and honey production, which sustain the livings of millions of coastal communities. They act as natural bio-shields, minimizing the economic losses from floods and storms, while simultaneously promoting ecotourism and offering avenues for sustainable coastal development, as shown in Figures 4 and 5.
Figure 4. Different economic significances of Mangrove ecosystems.
Figure 5. Mangrove ecosystems.
Fisheries Support
Mangroves serve as vital nursery grounds for commercially important fish and crustaceans, with their intricate root systems providing shelter from predators and currents for juvenile species (12, 63). Healthy mangrove forests directly enhance fishery productivity, as evidenced by positive correlations between mangrove presence and fish catch rates in adjacent waters (64). This productivity is highest in mangroves with robust growth, freshwater inputs, and intact ecosystems, particularly along fringe habitats where fish populations concentrate (65). The structural complexity of roots, tidal channels, and pools further supports fisheries by offering refuge and sustaining prey like bivalves (65). Beyond nursery functions, mangroves sustain food webs through organic matter export (e. g. , detritus) to seagrass beds and coral reefs, amplifying fishery yields (3). These ecosystem services underpin artisanal and commercial fisheries globally, supporting millions of livelihoods (6, 15). However, mangrove loss from aquaculture, deforestation, and urbanization has severely degraded fishery resources (66). Notably, while fish catches peak near human settlements where fishing effort and markets converge these mangroves face heightened threats from pollution and overfishing (65). Targeted conservation and sustainable management of these vulnerable areas are essential for securing both ecological integrity and long-term economic benefits.
Timber and Non-Timber Forest Products (NTFPs)
Mangrove forests ecosystem serve as vital reservoirs of both timber and non-timber forest products (NTFPs), supporting coastal communities through diverse economic and medicinal resources. The durable wood of Rhizophora, Avicennia, Bruguiera, Xylocarpus, and Sonneratia species is highly valued for its resistance to decay and damage by insects, making it ideal for construction (housing, boat building, furniture), fuelwood, and charcoal (7, 67-69). Beyond timber, mangroves provide nutritious fodder, particularly from Avicennia marina, sustaining livestocks (70). The medicinal potential of mangroves is equally significant, with traditional applications ranging from treating skin ailments and ulcers to dysentery (71, 68, 72). Bioactive compounds from species like Nypa fruticans and Ceriops spp. exhibit antimicrobial properties, while the mangrove-associated tunicate Ecteinascidia turbinata shows promise in cancer research (73, 74). Additionally, tannins from Rhizophora mucronata and Ceriops tagal are vital for leather tanning and dyeing (71). Mangroves further bolster local economies through NTFPs like honey (75) and fisheries (76), while their role in carbon sequestration and coastal protection highlights their ecological value (77, 55). However, overharvesting and habitat degradation threaten these ecosystems (78), necessitating community-based conservation and sustainable harvesting practices (8). Balancing utilization with preservation is key to ensuring mangroves continue to sustain livelihoods while maintaining their critical ecological functions.
Ecotourism and Recreation
Successful ecotourism models in Bali’s mangrove forestsand Vietnam’s Cần Giờ Biosphere Reserve demonstrate how tourism can be effectively integrated with biodiversity conservation (84, 85). Beyond recreation, mangrove ecotourism serves important educational functions. Research tourism, student field visits, and nature-based learning activities contribute to scientific understanding and promote environmental awareness (86). Health and wellness tourism, including meditation and therapeutic retreats, further utilize the calming and restorative qualities of mangrove environments (87). These diverse activities align closely with multiple Sustainable Development Goals particularly those related to responsible consumption, climate action, and marine ecosystem protection (88).
The economic contributions are substantial, with ecotourism generating employment, supporting small-scale enterprises, and providing alternative income streams for coastal communities (89). In regions such as the Sundarbans and the Everglades, tourism revenue directly supports conservation and management programs, reinforcing local incentives for ecosystem protection (79, 83). Overall, mangrove ecotourism represents a sustainable strategy for coastal zone management under increasing climatic and environmental pressures.
Water Filtration
Mangrove ecosystems function as highly efficient natural water purification systems in coastal environments. Their complex root networks trap sediments and contaminants (90), reducing offshore turbidity while offering protection against shoreline erosion (91). This filtration capacity safeguards adjacent habitats particularly coral reefs and seagrass meadows from sedimentation and nutrient overload, thus supporting the goals outlined in SDG 14: Life Below Water (92).
The saline environment in which mangroves thrive exemplifies their exceptional filtering capabilities. Some mangrove species can remove up to 90% of sodium ions from seawater (93) through a specialized multilayered root epidermis that selectively restricts Na⁺ entry (94). For instance, Rhizophora stylosa roots demonstrate a hierarchical pore structure with an elevated zeta potential in the outer layer, effectively excluding sodium ions and providing a model for sustainable desalination technologies (95).
In the Río Coco Solo estuary of Panama, mangrove vegetation significantly mitigates anthropogenic nutrient inputs (96-99). Many mangrove species also support phytostabilization of toxic metals within intertidal soils (99), reducing the mobility and ecological impact of heavy metals in polluted estuaries. These services are particularly crucial in regions experiencing high nutrient loads from agriculture, aquaculture, and urban wastewater.
Genetic Resources with Biotechnological Potential
Mangroves, adapted to extreme intertidal conditions, possess unique genetic and biochemical traits that enable them to withstand salinity, waterlogging, and oxidative stress. These specialized adaptations offer promising applications in biotechnology (100). Numerous mangrove-associated microorganisms such as Penicillium, Chaetomium, Humicola, Melanocarpus, and Bacillus species are utilized in diverse industrial sectors including pulp and paper processing, animal feed manufacturing, bakery and beverage fermentation, and xylitol production (101).
Marine algae such as Codium fragile, C. latum, C. dwarkense, and C. tomentosum produce compounds like phyllacoid, agar, carrageenan, and alginate, all demonstrating anticoagulant properties (102, 103). Fungal extracts containing ethyl acetate derivatives act as biopesticides (104, 105). Halophilic bacteria synthesize polyketide synthases (PKSs) responsible for producing medically important compounds such as erythromycin, rapamycin, tetracycline, lovastatin, and resveratrol (106).
Other enzymes including catalase, peroxidase, oxidase, polyphenol oxidase, and ascorbic acid oxidase from Pseudomonas aeruginosa, P. alcaligenes, and Methylococcus spp. hold significant industrial value (107). Additional enzymes such as proteases, amylases, lipases, and esterases produced by members of Vibronales, Actinomycetales, and Bacillales are used in food, pharmaceutical, and biofuel industries (108). Phytase derived from Bacillus circulans, B. licheniformis, and B. pantothenicus is important in agriculture (109), while lipopeptides and glycolipids from Bacillus subtilis and Pseudomonas aeruginosa act as effective biosurfactants (110).
Mangrove-associated fungi such as Preussia aurantiaca synthesize auranticins A and B with strong antimicrobial properties (111). Aigialus parvus yields aigialomycins A–E and hypothemicin, compounds with notable antimicrobial activity (112). Actinomycetes produce secondary metabolites exhibiting anti-cancer, anti-tumor, and anti-infective activities (113). Xylaria species generate xyloketals A–E, known inhibitors of acetylcholinesterase (114), while isoflavones and prostaglandin derivatives from Phomopsis, Paecilomyces, Sargassum, and Halorosellinia species display anti-cancer effects (115).
Bacteria such as Pseudomonas, Shewanella, Sphingomonas, Arthrobacter, Marinobacter, Alcanivorax, Microbulbifer, Micrococcus, Cellulomonas, Dietzia, and Gordonia contribute to bioremediation by degrading polyaromatic hydrocarbons and oil contaminants (116-121). Basidiomycetes enhance remediation efforts through the production of laccase (122, 123). Phosphatases from B. licheniformis, Chryseomonas luteola, Pseudomonas stutzeri, and Aspergillus niger provide agricultural benefits (124, 125), while halophilic archaea such as Halococcus synthesize L-asparaginase for industrial use (126). Enzymes like xylanase and ligninase, produced by Aspergillus niger and Phlebia spp. , are used in paper and biofuel industries (127). Biosurfactants from Leuconobacter, B. subtilis, and P. aeruginosa have wide biotechnological applications (128). Fusarium species release enniatin G, an antimicrobial compound (129), while Verruculina enalia produces enalin A and B with similar activities (114, 130-132). Additionally, fungal metabolites such as β-carboline, adenosine, and 8-hydroxy-3, 5-dimethylisochroman-1-one show potent anti-tumor properties (132). Stress-adapted genes from mangrove plants further provide valuable resources for developing crops tolerant to salinity and drought. NHX-type antiporters and osmoprotectant biosynthetic genes from Avicennia marina and Rhizophora species, when expressed in Arabidopsis, significantly improve stress tolerance (133). The microbial associates of the seagrass Syringodium isoetifolium showed notable antibacterial activity, with Actinomycetes sp. (Act01) emerging as the most potent isolate against antibiotic-resistant and fish bacterial pathogens. Phylogenetic and GC–MS analyses confirmed Act01 as a Streptomyces species producing a major anthraquinone compound, highlighting its potential as a novel antibacterial agent (134). Collectively, mangrove-associated microorganisms represent powerful reservoirs of novel bioactive metabolites and genetic resources with applications in pharmaceuticals, bioremediation, and climate-resilient agriculture (135).
A growing body of literature from 1991–2025 documents expanding mangrove research across Western Asia, with major themes centred on biodiversity, climate change adaptation, and socio-economic relevance (136). Reviews highlight key strengths such as the robustness of Avicennia marina and rising institutional collaboration (136). However, persistent weaknesses including fragmented governance and insufficient data integration are compounded by threats from climate change, industrial expansion, and transboundary pollution (136).
Research Gaps and Future Directions
Based on the synthesis of 135 studies, several cross-cutting research gaps emerge. First, empirical work is heavily concentrated in Southeast Asia, with limited data from Latin America, Africa, Western Asia, and small island states, underscoring the need for harmonized field and remote-sensing assessments in under-represented regions. Second, blue carbon research focuses largely on above-ground biomass and the top 1 m of soil, with deep soil carbon, lateral fluxes, and post-disturbance dynamics remaining poorly understood, calling for long-term, depth-resolved carbon studies. Third, economic valuations of fisheries, NTFPs, ecotourism, and carbon credits employ heterogeneous methods and rarely adopt standardized TEV or PES frameworks, limiting robust analysis of trade-offs and opportunity costs. Fourth, evidence on coastal protection, water filtration, and pollution control is often site-specific and short-term, highlighting the need for multi-site, long-term monitoring that links mangrove structure and water-quality regulation to risk-reduction metrics and downstream ecosystem health. Finally, biotechnological potential, governance, and carbon-credit schemes are underexplored beyond discovery-level studies; future research should move towards pre-clinical/industrial pipelines, rigorous analysis of governance and equity, robust MRV frameworks for blue carbon projects, and long-term evaluation of hydrologically sound restoration designs under climate-change scenarios.
Conclusion
Mangroves are among the planet’s most valuable yet increasingly threatened ecosystems. Although they occupy a relatively small portion of global coastlines, their ecological and socio-economic contributions are disproportionately large. Ecologically, mangroves function as biodiversity hotspots, nursery habitats for aquatic species, natural coastal defense systems, and highly efficient blue carbon stores substantially mitigating the impacts of climate change. Economically, they support millions of livelihoods through fisheries, timber and non-timber products, ecotourism, and emerging carbon-credit markets, while offering cost-effective alternatives to engineered coastal infrastructure. However, escalating pressures ranging from pollution and deforestation to aquaculture expansion and sea-level rise underscore the urgency of integrated conservation strategies. Protecting and restoring mangroves is both an ecological necessity and a socio-economic imperative, as their loss would jeopardize food security, coastal resilience, and climate regulation for millions of people. Future efforts must therefore prioritize ecosystem-based management, combining habitat restoration, community participation, robust policy frameworks, and sustainable resource use. Recognizing mangroves as both ecological keystones and economic assets will be essential for ensuring their continued role as nature-based solutions for a resilient and sustainable future.
Declarations
Conflict of Interest
The authors declare no conflicting interest.
Data Availability
The unpublished data is available upon request to the corresponding author.
Ethics Statement
Not applicable
Funding Information
The authors declare that no financial support was received for the research, authorship, and/or publication of this article.
References
Jennerjahn TC, Ittekot V. Relevance of mangroves for the production and deposition of organic matter along tropical continental margins. Naturwissenschaften. 2002; 89 (1): 23– 30.
Giri C, Ochieng E, Tieszen LL, Zhu Z, Singh A, Loveland T, Duke N. Status and distribution of mangrove forests of the world using earth observation satellite data. Glob Ecol Biogeogr. 2011; 20 (1): 154– 159.
Alongi DM. Present state and future of the world’ s mangrove forests. Environ Conserv. 2002; 29 (3): 331– 349.
Duke NC, Meynecke JO, Dittmann S, Ellison AM, Anger K, Berger U, Dahdouh-Guebas F. A world without mangroves? Science. 2007; 317 (5834): 41b– 42b.
Donato DC, Kauffman JB, Murdiyarso D, Kurnianto S, Stidham M, Kanninen M. Mangroves among the most carbon-rich forests in the tropics. Nat Geosci. 2011; 4 (5): 293– 297.
Mcleod E, Chmura GL, Bouillon S, Salm R, Bjö rk M, Duarte CM, Silliman BR. A blueprint for blue carbon: Toward an improved understanding of the role of vegetated coastal habitats in sequestering CO₂. Front Ecol Environ. 2011; 9 (10): 552– 560.
Barbier EB, Hacker SD, Kennedy C, Koch EW, Stier AC, Silliman BR. The value of estuarine and coastal ecosystem services. Ecol Monogr. 2011; 81 (2): 169– 193.
Walters BB, Rö nnbä ck P, Kovacs JM, Crona B, Hussain SA, Badola R, et al. Ethnobiology, socio-economics and management of mangrove forests: A review. Aquat Bot. 2008; 89 (2): 220– 236.
Valiela I, Bowen JL, York JK. Mangrove forests: One of the world's threatened major tropical environments. BioScience. 2001; 51 (10): 807– 815.
Moher D, Liberati A, Tetzlaff J, Altman DG. Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement. PLoS Med. 2009; 6: e1000097. doi: 10.1371/journal. pmed.1000097.
Hogarth PJ. The Biology of Mangroves and Seagrasses. 3rd ed. Oxford: Oxford University Press; 2015.
Nagelkerken I, Blaber SJM, Bouillon S, Green P, Haywood M, Kirton LG, et al. The habitat function of mangroves for terrestrial and marine fauna: A review. Aquat Bot. 2008; 89 (2): 155– 185.
Laegdsgaard P, Johnson C. Why do juvenile fish utilise mangrove habitats? J Exp Mar Biol Ecol. 2001; 257 (2): 229– 253.
Mumby PJ, Edwards AJ, Arias-Gonzá lez JE, Lindeman KC, Blackwell PG, Gall A, Llewellyn G. Mangroves enhance the biomass of coral reef fish communities in the Caribbean. Nature. 2004; 427 (6974): 533– 536.
Aburto-Oropeza O, Ezcurra E, Danemann G, Valdez V, Murray J, Sala E. Mangroves in the Gulf of California increase fishery yields. Proc Natl Acad Sci U S A. 2008; 105 (30): 10456– 10459.
Nusantara MA, Malikusworo H, Helmi P. Evaluation and planning of mangrove restoration programs in Sedari Village of Kerawang District, West Java: Coastal development programs. Procedia Environ Sci. 2015; 23: 207– 214.
Adi W, Sari SP. Detection of mangrove distribution in Pongok Island. Procedia Environ Sci. 2016; 33: 253– 257.
Spalding M, Kainuma M, Collins L. World Atlas of Mangroves. London: Earthscan; 2010.
Luther DA, Greenberg R. Mangroves: A global perspective on the evolution and conservation of their terrestrial vertebrates. BioScience. 2009; 59 (7): 602– 612.
Hattam C, Goh HC, Then AY, Edwards-Jones A, Ruslan NF, Yap JS, Moh HH. Using nexus thinking to identify opportunities for mangrove management in the Klang Islands, Malaysia. Estuar Coast Shelf Sci. 2021; 248: 107157.
Romañ ach SS, DeAngelis DL, Koh HL, Li Y, Teh SY, Barizan RSR, Zhai L. Conservation and restoration of mangroves: Global status, perspectives, and prognosis. Ocean Coast Manag. 2018; 154: 72– 82.
Cannicci S, Burrows D, Fratini S, Smith III TJ, Offenberg J, Dahdouh-Guebas F. Faunal impact on vegetation structure and ecosystem function in mangrove forests: A review. Aquat Bot. 2008; 89 (2): 186– 200.
Macdonald DW, Newman C, Harrington LA. The biology and conservation of wild canids. In: Macdonald DW, Loveridge AJ, editors. Biology and Conservation of Wild Felids. Oxford: Oxford University Press; 2010. p.139– 160.
Pasquaud S, Vasconcelos RP, Franç a S, Henriques S, Costa MJ, Cabral H. Worldwide patterns of fish biodiversity in estuaries: Effect of global vs local factors. Estuar Coast Shelf Sci. 2015; 154: 122– 128.
Suratman MN. Carbon sequestration potential of mangroves in Southeast Asia. In: Bravo F, LeMay V, Jandl R, von Gadow K, editors. Managing Forest Ecosystems: The Challenge of Climate Change. Dordrecht: Springer; 2008. p.297– 315.
McIvor AL, Mö ller I, Spencer T, Spalding M. Reduction of wind and swell waves by mangroves. Natural Coastal Protection Series Report 1. The Nature Conservancy and Wetlands International; 2012.
McIvor AL, Mö ller I, Spencer T, Spalding M. Storm surge reduction by mangroves. Natural Coastal Protection Series Report 2. The Nature Conservancy and Wetlands International; 2012.
Mazda Y, Magi M, Kogo M, Hong PN. Mangroves as a coastal protection from waves in the Tong King delta, Vietnam. Mangroves Salt Marshes. 1997; 1 (2): 127– 135.
Zhang KQ, Liu H, Li Y, Hongzhou X, Jian S, Rhome J, Smith III TJ. The role of mangroves in attenuating storm surges. Estuar Coast Shelf Sci. 2012; 102: 11– 23.
Liu H, Zhang K, Li Y, Xie L. Numerical study of the sensitivity of mangroves in reducing storm surge and flooding to hurricane characteristics in southern Florida. Cont Shelf Res. 2013; 64: 51– 65.
Yanagisawa H, Koshimura S, Miyagi T, Imamura F. Tsunami damage reduction performance of a mangrove forest in Banda Aceh, Indonesia inferred from field data and a numerical model. J Geophys Res Oceans. 2010; 115 (C6).
Furukawa K, Wolanski E. Sedimentation in mangrove forest. Mangroves Salt Marshes. 1996; 1 (1): 3– 10.
Krauss KW, McKee KL, Lovelock CE, Cahoon DR, Saintilan N, Reef R, Chen L. How mangrove forests adjust to rising sea level. New Phytol. 2014; 202 (1): 19– 34.
Lovelock CE, Cahoon DR, Friess DA, Guntenspergen GR, Krauss KW, Reef R, Saintilan N. The vulnerability of Indo-Pacific mangrove forests to sea-level rise. Nature. 2015; 526 (7574): 559– 563.
Winterwerp H, Wesenbeeck BV, Dalfsen JV, Tonneijck F, Astra A, Verschure S, Eijk PV. A sustainable solution for massive coastal erosion in Central Java: Towards regional scale application of hybrid engineering. Discussion Paper. 2014.
Asari N, Suratman MN, Ayob N, Hamid N. Mangrove as a natural barrier to environmental risks and coastal protection. In: Mangroves: Ecology, Biodiversity and Management. Springer; 2021. p.1– 20.
Chang SE, Adams BJ, Alder J, Berke PR, Chuenpagdee R, Ghosh S, Wabnitz C. Coastal ecosystems and tsunami protection after the December 2004 Indian Ocean tsunami. Earthq Spectra. 2006; 22 (S3): 863– 887.
Knutson TR, McBride JL, Chan J, Emanuel K, Holland G, Landsea C, et al. Tropical cyclones and climate change. Nat Geosci. 2010; 3 (3): 157– 163.
Alongi DM. Carbon cycling and storage in mangrove forests. Annu Rev Mar Sci. 2014; 6: 195– 219.
Bastos RF, Lippi DL, Gaspar ALB, Yogui GT, Fré dou T, Garcia AM, Ferreira BP. Ontogeny drives allochthonous trophic support of snappers: Seascape connectivity along the mangrove– seagrass– coral reef continuum of a tropical marine protected area. Estuar Coast Shelf Sci. 2022; 264: 107591.
Mishra AK, Apte D. Ecological connectivity with mangroves influences tropical seagrass population longevity and meadow traits within an island ecosystem. Mar Ecol Prog Ser. 2020; 644: 47– 63.
Watanabe A, Nakamura T. Carbon dynamics in coral reefs. In: Blue Carbon in Shallow Coastal Ecosystems: Carbon Dynamics, Policy, and Implementation. Springer; 2019. p.273– 293.
Akhand A, Watanabe K, Chanda A, Tokoro T, Chakraborty K, Moki H, et al. Lateral carbon fluxes and CO₂ evasion from a subtropical mangrove– seagrass– coral continuum. Sci Total Environ. 2021; 752: 142190.
Lovelock CE, Reef R. Variable impacts of climate change on blue carbon. One Earth. 2020; 3 (2): 195– 211.
Lee SY. Mangrove outwelling: A review. Hydrobiologia. 1995; 295 (1– 3): 203– 212.
Fusi M, Booth JM, Marasco R, Merlino G, Garcias-Bonet N, Barozzi A, et al. Bioturbation intensity modifies the sediment microbiome and biochemistry and supports plant growth in an arid mangrove system. Microbiol Spectr. 2022; 10: e01117– e01122.
Kristensen E, Bouillon S, Dittmar T, Marchand C. Organic carbon dynamics in mangrove ecosystems: A review. Aquat Bot. 2008; 89 (2): 201– 219.
Bouillon S, Borges AV, Castañ eda-Moya E, Diele K, Dittmar T, Duke NC, et al. Mangrove production and carbon sinks: A revision of global budget estimates. Glob Biogeochem Cycles. 2008; 22 (2).
Tam NFY, Wong YS. Mangrove soils as sinks for wastewater-borne pollutants. Hydrobiologia. 1995; 295 (1– 3): 231– 241.
Shaltout KH, Ahmed MY, Alrumman SA, Ahmed DA, Eid EM. Evaluation of the carbon sequestration capacity of arid mangroves along nutrient availability and salinity gradients along the Red Sea coastline of Saudi Arabia. Oceanologia. 2020; 62 (1): 56– 69.
Murdiyarso D, Donato D, Kauffman JB, Kurnianto S, Stidham M, Kanninen M. Carbon storage in mangrove and peatland ecosystems: A preliminary account from plots in Indonesia. Working Paper 48. Center for International Forestry Research (CIFOR); 2009. p.39.
Trumper K, Bertzky M, Dickson B, van der Heijden G, Jenkins M, Manning P. The natural fix? The role of ecosystems in climate mitigation. UNEP Rapid Response Assessment. UNEP– WCMC, Cambridge, UK; 2009. p.65.
Eid EM, Shaltout KH, Al-Sodany YM, Haroun SA, Galal TM, Ayed H, Khedher KM. Carbon sequestration potential of mangrove soils in the Nile Delta, Egypt. Glob Ecol Conserv. 2020; 24: e01390.
Friess DA. Ecosystem services and disservices of mangrove forests. Curr For Rep. 2016; 2 (1): 30– 41.
Askari M, Homaei A, Kamrani E, Zeinali F, Andreetta A. Estimation of carbon pools in the biomass and soil of mangrove forests in Sirik Azini creek, Hormozgan province (Iran). Environ Sci Pollut Res. 2022; 29: 23712– 23720.
Song S, Ding Y, Li W, Meng Y, Zhou J, Gou R, et al. Mangrove reforestation provides greater blue carbon benefit than afforestation for mitigating global climate change. Nat Commun. 2023; 14: 756.
Zhu JJ, Yan B. Blue carbon sink function and carbon neutrality potential of mangroves. Sci Total Environ. 2022; 822: 153438.
Leal M, Spalding MD. The State of the World’ s Mangroves 2022. Global Mangrove Alliance; 2022.
Sumarmi S, Arinta D, Suprianto A, Aliman M. The development of ecotourism with community-based tourism (CBT) in Clungup Mangrove Conservation (CMC) of Tiga Warna Beach for sustainable conservation. Folia Geogr. 2021; 63 (1): 123– 138.
Bunting P, Rosenqvist A, Hilarides L, Lucas RM, Thomas N. Global mangrove extent change 1996– 2020: Global Mangrove Watch version 3.0. Remote Sens. 2022; 14 (15): 3657.
Alsumaiti TS, Shahid SA. Mangroves among most carbon-rich ecosystems living in hostile saline-rich environments and mitigating climate change— A case of Abu Dhabi. J Agric Crop Res. 2019; 7 (1): 1– 8.
Manson FJ, Loneragan NR, Harch BD, Skilleter GA, Williams L. A broad-scale analysis of links between coastal fisheries production and mangrove extent: A case study for northeastern Australia. Fish Res. 2005; 74 (1– 3): 69– 85.
Pauly D, Ingles J. The relationship between shrimp yields and intertidal vegetation (mangrove) areas: A reassessment. In: IOC/FAO Workshop on Recruitment in Tropical Coastal Demersal Communities. Ciudad del Carmen, Mexico; 1986.
Hutchison J, Spalding M, zu Ermgassen P. The Role of Mangroves in Fisheries Enhancement. The Nature Conservancy and Wetlands International; 2014. 54 p.
Primavera JH. Mangroves, fishponds, and the quest for sustainability. Science. 2005; 310 (5745): 57– 59.
Kairo JG, Dahdouh-Guebas F, Bosire J, Koedam N. Restoration and management of mangrove systems— A lesson for and from the East African region. S Afr J Bot. 2001; 67 (3): 383– 389.
Kusmana C. Mangrove plant utilization by local coastal community in Indonesia. IOP Conf Ser Earth Environ Sci. 2017; 126 (1): 012028.
Olorunnisola AO. The past, present and future outlook of the wood industry in Nigeria. In: Sustainable Forest Management. IntechOpen; 2023. p.1– 20.
Mitra A. Ecosystem services of mangroves: An overview. In: Mitra A, editor. Mangrove Forests in India: Exploring Ecosystem Services. Cham: Springer; 2020. p.1– 32.
Bandaranayake WM. Traditional and medicinal uses of mangroves. Mangroves Salt Marshes. 1998; 2 (3): 133– 148.
Friess DA, Yando ES, Alemu JB, Wong L-W, Soto SD, Bhatia N, et al. Ecosystem services and disservices of mangrove forests and salt marshes. Oceanogr Mar Biol Annu Rev. 2020; 58: 107– 142.
Vinoth R, Kumaravel S, Ranganathan R. Therapeutic and traditional uses of mangrove plants. J Drug Deliv Ther. 2019; 9 (4): 849– 854.
Habib MA, Khatun F, Ruma M, Chowdhury A, Silve A, Rahman A, et al. A review on phytochemical constituents of pharmaceutically important mangrove plants, their medicinal uses and pharmacological activities. Vedic Res Int Phytomed. 2018; 6 (1): 1– 9.
Krauss KW, Doyle TW, Doyle TJ, Swarzenski CM, From AS, Day RH, Conner WH. Water level observations in mangrove swamps during two hurricanes in Florida. Wetlands. 2009; 29 (1): 142– 149.
Krauss KW, Doyle TW, Twilley RR, Smith TJ, Whelan KRT, Sullivan JK. Sea-level rise and landscape change influence mangrove encroachment onto marsh in the Ten Thousand Islands region of Florida, USA. J Coast Conserv. 2009; 13 (1): 1– 11.
Anneboina LR, Kumar KV. Economic analysis of mangrove and marine fishery linkages in India. Ecosyst Serv. 2017; 24: 114– 123.
Dahdouh-Guebas F, Hettiarachchi S, Sooriyarachchi S, Lo Seen D, Batelaan O, Jayatissa LP, Koedam N. Transitions in ancient inland freshwater resource management in Sri Lanka affect biota and human populations in and around coastal lagoons. Curr Biol. 2005; 15: 579– 58
Badola R, Barthwal S, Hussain SA. Attitudes of local communities towards conservation of mangrove forests: A case study from the east coast of India. Estuar Coast Shelf Sci. 2012; 96 (4): 423– 432.
Satyanarayana B, Bhanderi P, Debry M, Maniatis D, Foré F, Badgie D, et al. A socio-ecological assessment aiming at improved forest resource management and sustainable ecotourism development in the mangroves of Tanbi Wetland National Park, The Gambia, West Africa. Ambio. 2012; 41 (5): 513– 526.
Uddin MS, van Steveninck EdR, Stuip M, Shah MAR. Economic valuation of provisioning and cultural services of a protected mangrove ecosystem: A case study on Sundarbans Reserve Forest, Bangladesh. Ecosyst Serv. 2013; 5: 88– 93.
Basyuni M, Bimantara Y, Siagian M, Wati R, Slamet B, Sulistiyono N, et al. Developing community-based mangrove management through ecotourism in North Sumatra, Indonesia. IOP Conf Ser Earth Environ Sci. 2017; 126: 012109.
Gumede TK, Nzama AT. Approaches toward community participation enhancement in ecotourism. Protected Area Management – Recent Advances. 2022; 1 (1): 1– 13.
Gaora PA, Pedrason R, Herman E. Indonesia’ s climate diplomacy under Joko Widodo: Shaping equitable and sustainable global future. Nation State J Int Stud. 2023; 6 (1): 34– 48.
Spalding M, McIvor A, Tonneijck FH, Tol S, van Eijk P. Mangroves for Coastal Defence: Guidelines for Coastal Managers & Policy Makers. Wetlands International and The Nature Conservancy; 2014.
Avau J, Cunha-Lignon M, De Myttenaere B, Godart M-F, Dahdouh-Guebas F. The commercial images promoting Caribbean mangroves to tourists: Case studies in Jamaica, Guadeloupe and Martinique. J Coast Res. 2011; 64: 1277– 1281.
Khoshtaria T, Chachava N. Prospects of ecotourism development in recreation areas of South Georgia. Ann Agrar Sci. 2017; 15 (3): 312– 317.
Das S. Ecotourism, sustainable development and the Indian state. Econ Polit Wkly. 2011; 46 (2): 60– 67.
Ghosh S, Ghosh A, Choudhury SB. Mangrove forest and ecotourism: A case study of the Sundarbans. Int J Sci Res Publ. 2015; 5 (4): 1– 6.
Sultana S, Huang R, Van Zwieten L, Wang H, Wu J. Trapping effect of mangrove and saltmarsh habitats on geochemical elements: A case study in Ximen Island, Zhejiang, China. J Soils Sediments. 2023; 23 (5): 2327– 2343.
Maza M, Lara JL, Losada IJ. Predicting the evolution of coastal protection service with mangrove forest age. Coast Eng. 2021; 168: 103922.
Liu C, Liu G, Yang Q, Luo T, He P, Franzese PP, Lombardi GV. Emergy-based evaluation of world coastal ecosystem services. Water Res. 2021; 204: 117656.
Hastuti ED, Izzati M, Prihastanti E. Water uptake and salt accumulation under Rhizophora stylosa seedling planted in controlled salinity and inundation levels. AACL Bioflux. 2023; 16 (2): 1069– 1076.
Li X, Kondo R, Sakai K. Biodegradation of sugarcane bagasse with marine fungus Phlebia sp. MG– 60. J Wood Sci. 2002; 48 (2): 159– 162.
Kim K, Seo E, Chang SK, Park TJ, Lee SJ. Novel water filtration of saline water in the outermost layer of mangrove roots. Sci Rep. 2016; 6: 20426.
Jusoff K. Malaysian mangrove forests and their significance to the coastal marine environment. Pol J Environ Stud. 2013; 22 (4): 979– 1005.
Wang Q, Mei D, Chen J, Lin Y, Liu J, Lu H, Yan C. Sequestration of heavy metal by glomalin-related soil protein: Implication for water quality improvement in mangrove wetlands. Water Res. 2019; 148: 142– 152.
Lin B, Dushoff J. Mangrove filtration of anthropogenic nutrients in the Rio Coco Solo, Panama. Manag Environ Qual. 2004; 15 (2): 131– 142.
Zhou YW, Peng YS, Li XL, Chen GZ. Accumulation and partitioning of heavy metals in mangrove rhizosphere sediments. Environ Earth Sci. 2011; 64 (3): 799– 807.
Alongi DM. The impact of climate change on mangrove forests. Curr Clim Change Rep. 2015; 1 (1): 30– 39.
Polizeli MLTM, Rizzatti ACS, Monti R, Terenzi HF, Jorge JA, Amorim DS. Xylanases from fungi: properties and industrial applications. Appl Microbiol Biotechnol. 2005; 67 (5): 577– 591.
Shanmugam S, Mody KH. Heparinoid active sulphated polysaccharides from marine algae as potential blood coagulant agents. Curr Sci. 2000; 79 (12): 1672– 1683.
Xiao YT, Zheng ZH, Huang YJ, Xu QY, Su WJ, Song SY. Nematicidal and brine shrimp lethality of secondary metabolites from marine-derived fungi. J Xiamen Univ Nat Sci. 2005; 44 (6): 847– 850.
Chen GY, Lin YC, Wen L, Vrijmoed LLP, Jones EBG. Two new metabolites of a marine endophytic fungus from an estuarine mangrove on the South China Sea coast. Tetrahedron. 2003; 59 (26): 4907– 4909.
Ventosa A, Nieto JJ. Biotechnological applications and potentialities of halophilic microorganisms. World J Microbiol Biotechnol. 1995; 11 (1): 85– 94.
Mishra RR. Microbial biodiversity in mangroves of Bhitarakanika, Orissa— a study on genotypic, phenotypic and proteomic characterisation of some predominant bacteria. PhD Thesis. North Orissa University; 2010.
Armando CFD, Andreote FD, Dini-Andreote F, Lacava PT, Sá ALB, Melo IS. Diversity and biotechnological potential of culturable bacteria from Brazilian mangrove sediment. World J Microbiol Biotechnol. 2009; 25: 1305– 1311.
Joseph I, Paul Raj R. Isolation and characterization of phytase producing Bacillus strains from mangrove ecosystem. J Mar Biol Assoc India. 2007; 49 (2): 177– 182.
Pornsunthorntawee O, Wongpanit P, Chavadej S, Abe M, Rujiravanit R. Structural and physicochemical characterization of crude biosurfactants produced by Pseudomonas aeruginosa SP4 isolated from petroleum-contaminated soil. Bioresour Technol. 2008; 99 (6): 1589– 1595.
Poch GK, Gloer JB. Helicascolides A and B: new lactones from the marine fungus Helicascus kunaloanus. J Nat Prod. 1989; 52 (2): 257– 260.
Isaka M, Suyarnsestakorn C, Tanticharoen M. Aigialomycins A– E, new resorcylic macrolides from the marine mangrove fungus Aigialus parvus. J Org Chem. 2002; 67 (5): 1561– 1566.
Hong K, Gao AH, Xie QY, Gao H, Zhuang L, Lin HP, et al. Actinomycetes for marine drug discovery isolated from mangrove soils and plants in China. Mar Drugs. 2009; 7: 24– 4.
Lin YC, Wu XY, Feng S, Jiang GC, Luo JH, Zhou SN, et al. Five unique compounds: xyloketals from mangrove fungus Xylaria sp. from the South China Sea coast. J Org Chem. 2001; 66 (19): 6252– 6256.
Tao L, Zhang JY, Liang YJ, Chen LM, Zhen LC, Wang F, et al. Anticancer effect and structure– activity analysis of marine products isolated from metabolites of mangrove fungi in the South China Sea. Mar Drugs. 2010; 8 (4): 1094– 1105.
Desai C, Pathak H, Madamwar D. Advances in molecular and “ omics” technologies to gauge microbial communities and bioremediation at xenobiotic/anthropogen contaminated sites. Bioresour Technol. 2010; 101 (6): 1558– 1569.
Brito EM, Guyoneaud R, Goñ i-Urriza M, Ranchou-Peyruse A, Verbaere A, Crapez MAC. Characterization of hydrocarbonoclastic bacterial communities from mangrove sediments in Guanabara Bay, Brazil. Res Microbiol. 2006; 157 (8): 752– 762.
Ramsay MA, Swannell RPJ, Shipton WA, Duke NC, Hill RT. Effect of bioremediation community in oiled mangrove sediments. Mar Pollut Bull. 2000; 41 (7– 12): 413– 419.
Yu KS, Wong AH, Yau KW, Wong YS, Tam NF. Natural attenuation, biostimulation and bioaugmentation on biodegradation of polycyclic aromatic hydrocarbons (PAHs) in mangrove sediments. Mar Pollut Bull. 2005; 51 (8– 12): 1071– 1077.
Ke L, Wang WQ, Wong TWY, Wong YS, Tam NFY. Removal of pyrene from contaminated sediments by mangrove microcosms. Chemosphere. 2003; 51 (1): 25– 34.
Mishra RR, Prajapati S, Das J, Dangar TK, Das N, Thatoi HN. Reduction of selenite to red elemental selenium by moderately halotolerant Bacillus megaterium strains isolated from Bhitarkanika mangrove soil and characterization of reduced product. Chemosphere. 2011; 84 (9): 1231– 1237.
D’ Souza DT, Tiwari R, Sah AK, Raghukumar C. Enhanced production of laccase by a marine fungus during treatment of colored effluents and synthetic dyes. Enzyme Microb Technol. 2006; 38: 504– 511.
Sabu A. Sources, properties and applications of microbial therapeutic enzymes. Indian J Biotechnol. 2003; 2 (3): 334– 341.
Vazquez P, Holguin G, Puerte ME, Lopez-Cortes A, Bashan Y. Phosphate-solubilising microorganisms associated with the rhizosphere of mangroves in a semiarid coastal lagoon. Biol Fertil Soils. 2000; 30 (5): 460– 468.
Gyaneshwar P, Kumar GN, Parekh LJ. Effect of buffering on the P-solubilizing ability of microorganisms. World J Microbiol Biotechnol. 1998; 14 (5): 669– 673.
Sudha V. Studies on halophilic bacteria associated with mangrove sediment and a biovalve mollusc Anadara rhombea (Born) (Arcidae) and on L-asparaginase (Anti-Leukamic agent) from an extremely halophilic bacterium. PhD Thesis. Annamalai University; 1981.
Saimmai A, Sobhon V, Maneerat S. Production of biosurfactants from a new and promising strain of Leucobacter komagatae 183. Ann Microbiol. 2011; 62 (1): 391– 402.
Lin YC, Wang J, Wu XY, Zhou SN, Vrijmoed LLP, Jones EBG. A novel compound enniatin G from the mangrove fungus Halosarpheia sp. (strain 732) from the South China Sea. Aust J Chem. 2002; 55 (3): 225– 227.
Lin YC, Wu XY, Deng ZJ, Wang J, Zhou SN, Vrijmoed LLP, Jones EBG. The metabolites of the mangrove fungus Verruculina enalia No. 2606 from a salt lake in the Bahamas. Phytochemistry. 2002; 59 (5): 469– 471.
Huang H, Feng X, Xiao Z, Liu L, Li H, Ma L, Lin Y. Azaphilones and p-terphenyls from the mangrove endophytic fungus Penicillium chermesinum (ZH4-E2) isolated from the South China Sea. J Nat Prod. 2011; 74 (5): 997– 1002.
Xu MJ, Gessner G, Groth I, Lange C, Christner A, Bruhn T, Lin WH. Shearing D– K, new indole triterpenoids from an endophytic Penicillium sp. (strain HKI0459) with blocking activity on large-conductance calcium-activated potassium channels. Tetrahedron. 2007; 63 (2): 435– 444.
Song XH, Liu XH, Lin YC. Metabolites of mangrove fungus No. K23 and interaction of carboline with DNA. J Trop Oceanogr. 2004; 23 (3): 66– 71.
Jithesh MN, Prashanth SR, Sivaprakash KR, Parida AK. Antioxidative response mechanisms in halophytes: Their role in stress defence. J Genet. 2006; 85 (3): 237– 254.
Ravikumar S, Gnanadesigan M, Saravanan A, Monisha N, Brindha V, Muthumari S. Antagonistic properties of seagrass-associated Streptomyces sp. RAUACT-1: A source for anthraquinone-rich compound. Asian Pac J Trop Med. 2012; 5 (11): 887– 890.
Paul S, Siddiqua F, Tayung K. Exploration of mangroves associated microbes for bioactive metabolites. Int J Plant Environ. 2023; 9: 202– 209.
Yap Chee Kong, Al-Mutairi KA. Mangrove ecosystems in Western Asia: A literature review of trends, conservation gaps, and sustainable management strategies. Front For Glob Change. 2025; 8.