RESEARCH ARTICLE
Prevalence and Intensity of Endoparasites in Ribbonfish (Trichiurus lepturus) from (Lamongan) and Tamperan (Pacitan) Fish Landing Sites (TPI)
Aquatic Life Sciences|Vol. 3, Issue 1, pp. 1-8 (2026)
CC BY 4.0-2026 Authors
Views
Downloads
Shares
Received
Sep 29, 2025Revised
Dec 4, 2025Accepted
Dec 9, 2025Published
Jan 5, 2026
Abstract
Parasitic infections in marine fish pose economic and public health risks, particularly when zoonotic species are involved. Ribbonfish (Trichiurus lepturus), a commercially important species in Indonesia, is highly susceptible to endoparasites that may reduce fish quality and threaten human consumers. This study aimed to identify the types, prevalence, and intensity of endoparasites infecting ribbonfish landed at Paciran (Lamongan) and Tamperan (Pacitan). A total of 56 fish samples were examined through necropsy and microscopic observation of internal organs. Two nematode species, Anisakis sp. and Camallanus sp., were detected in the intestine, stomach, and muscle tissues. Prevalence was 0.2% at Paciran and 0.4% at Tamperan, while intensity reached 1.3 and 1.9 ind/individual, respectively. Infected fish were typically larger, with body weights ranging from 100–400 g and lengths of 60–80 cm, and often contained undigested small fish and crustaceans that may serve as intermediate hosts. Although infection levels were classified as low, the detection of zoonotic Anisakis sp. underscores the need for continued monitoring to prevent potential health risks and safeguard fishery resources.
Keywords:
Introduction
Parasitic infections in marine fish pose significant threats to both fisheries sustainability and public health. Ribbonfish (Trichiuridae), a commercially valuable demersal species widely distributed in Indonesian waters, represents an important fishery commodity for domestic consumption and international trade (1). However, endoparasites such as Anisakis spp. and Camallanus spp. are frequently reported in demersal fish, including ribbonfish, and can compromise fish quality, reduce market value, and act as potential zoonotic agents (2). Previous studies have reported high prevalence rates of parasitic infections in marine fishes across several Indonesian fish landing sites, underscoring the urgent need for systematic monitoring of fish health status to ensure product safety and sustainability (3).
The increasing global demand for ribbonfish, particularly from East Asian countries, has heightened the need for stringent quality control measures (4). According to the Indonesian Ministry of Marine Affairs and Fisheries (KKP), fishery products exported internationally must be accompanied by a health certificate (SKIPP/Health Certificate for Fish and Fish Products) confirming they are free from quarantine-related pathogenic infections, in compliance with international quality and safety standards. Despite these regulations, comprehensive data on the prevalence and intensity of endoparasite infections in ribbonfish from key fish landing sites in Indonesia remain limited (5). Moreover, infection dynamics may vary according to ecological conditions, fishing grounds, and handling practices, creating additional challenges in disease prevention and control. The lack of systematic surveillance may expose consumers to health risks and fishers to economic losses.
To address this gap, the present study investigates the prevalence and intensity of endoparasites in ribbonfish landed at Fish Landing Sites (TPI), namely Paciran in Lamongan (Java Sea) and Tamperan in Pacitan (Indian Ocean). By employing purposive sampling and necropsy examination of internal organs, this study provides baseline epidemiological data on endoparasite infection in ribbonfish from two ecologically distinct fishing grounds. The findings are expected to contribute to improved fish health management, food safety assurance, and the sustainable utilization of marine resources.
Methodology
Study Design and Rationale
This study employed a cross-sectional survey design to determine the prevalence and intensity of endoparasite infections in ribbonfish landed at Fish Landing Sites (TPI) in Paciran, Lamongan (Java Sea) and Tamperan, Pacitan (Indian Ocean). These two sites were selected because they represent ecologically distinct fishing grounds that allow comparison of infection dynamics under different marine environments.
Sampling Strategy and Population
Fish samples were collected between May and July 2022 through purposive sampling. Random sampling was not feasible because ribbonfish were landed in mixed catches and were not separated by species during auction transactions, so random withdrawal would have interfered with the commercial selling process. Approximately 10% of the daily ribbonfish catch landed at each TPI was obtained, resulting in a total of 56 specimens (n = 27 from Paciran and n = 29 from Tamperan). Only freshly landed individuals free from visible physical damage were included. Total body length (cm) and weight (g) were measured prior to necropsy.
Materials and Examination Procedures
Parasitological examinations were conducted at the Integrated Laboratory of Universitas Islam Negeri Sunan Ampel Surabaya. Dissection instruments (scalpels, scissors, forceps), Petri dishes, and compound microscopes (40×–400×) were used. Each specimen was dissected aseptically, and internal organs including the stomach, intestines, liver, kidneys and muscle tissues were examined for endoparasites.
Tissue digestion was conducted using 0.5% pepsin-HCl solution at 37 °C for 2–4 h; fluorescence screening was carried out using UV illumination at 365 nm to detect larvae inside tissues; larval extraction was performed using saline sedimentation followed by stereomicroscopic observation. Parasites were preserved in 70% ethanol and identified morphologically using standard taxonomic identification keys.
Variables and Measurements
Prevalence was calculated as the proportion of infected fish relative to the total number of examined fish (%). Infection intensity was defined as the mean number of parasites per infected fish. When multiple parasite species were found in a single host, the prevalence of each species was calculated separately based on species-specific parasite counts rather than pooled parasite totals to avoid overestimation.
Data Analysis
Data were analyzed descriptively and expressed as prevalence (%) and mean intensity (individuals per infected fish). Comparisons between sampling locations were performed using chi-square tests for prevalence and Mann–Whitney U tests for infection intensity. All statistical outputs including p-values, effect sizes, and 95% confidence intervals—are now reported in the Results section. The significance level was set at p < 0.05 using SPSS software (IBM Corp. , Armonk, NY, USA).
Ethical Considerations
All procedures followed institutional guidelines for animal handling and welfare. No fish were intentionally captured from the wild for the purpose of this study; all specimens were obtained exclusively from routine commercial landings, and no additional harm was inflicted beyond necropsy. Under Indonesian research regulations, post-mortem parasitological examination of commercially landed fish does not require an environmental or wildlife sampling permit; therefore, no permit number applies.
Results and Discussion
Fish Sampling
Fish sampling was conducted at two major landing sites representing distinct ecological regions: Bawean Bay, Gresik (Java Sea), for specimens landed at Paciran Fish Landing Site (TPI), and Prigi Bay, Trenggalek (Indian Ocean), for specimens landed at Tamperan TPI. Both areas fall within the Indonesian Fisheries Management Areas and are known as productive coastal ecosystems where ribbonfish (Trichiuridae), together with various demersal and pelagic fishes, are commonly landed (6).
Paciran Fish Landing Site (Lamongan)
Fishermen operating at the Fish Landing Site (TPI), Lamongan, generally conduct daily fishing trips in nearby waters, with ribbonfish (Trichiuridae) often caught as bycatch in Bawean Bay. During non-peak seasons, catches of ribbonfish per vessel range between 5–10 kg, whereas in peak seasons yields can reach up to 50 kg per trip. In this study, a total of 36 individuals (≈5 kg) were sampled, with body weights predominantly in the 100–200 g class and total lengths mostly exceeding 50 cm, consistent with the length and weight distributions presented in Table 1.
| Characteristics | Category | Average Number of Fish | |
|---|---|---|---|
| Paciran (Java Sea) | Tamperan (Indian Ocean) | ||
| Length Class (cm) | < 50 | 45.31 ± 4.09 | 67.57 ± 1.98 |
| > 50 | 57.27 ± 5.54 | 73.42 ± 2.73 | |
| Weight Class (g) | < 100 | 80.95 ± 15.52 | 185.70 ± 15.98 |
| 100–200 | 128.20 ± 27.75 | 245.93 ± 24.30 | |
| > 200 | 205.00 ± 7.07 | 335.30 ± 23.62 | |
Besides ribbonfish, other landed species included small pelagic fishes such as sardines (Sardinella spp. ), mackerel (Scomberomorus spp. ), tuna (Thunnus spp. ), skipjack (Katsuwonuspelamis), and scads (Decapterus spp. ), as well as crustaceans including crabs, swimming crabs, shrimp, shellfish, and squids (7). According to Fisheries Management Area (WPP NRI) 712, which covers the Java Sea, major fishing grounds are located around the Karimunjawa Islands, Bawean Island, Masalembu Islands, and the eastern parts of Matasiri Island. This region is highly productive, supporting small and large pelagic fish, demersal fish, and reef fish, a condition that also provides diverse potential intermediate and paratenic hosts for fish parasites (6).
The Bawean Bay fishing ground, where ribbonfish landed at Paciran are caught, is a productive coastal area characterized by sandy–muddy substrates and relatively shallow waters (< 50 m), which provide suitable habitat for demersal species, including Trichiuridae. The region supports diverse assemblages of pelagic and demersal fishes and generally meets national seawater quality standards, which may influence the structure of local fish communities and parasite transmission dynamics (6-9).
Tamperan Fish Landing Site (Pacitan)
At the Tamperan Fish Landing Site (TPI), Pacitan, ribbonfish (Trichiuridae) are harvested using both small and medium-sized vessels. During the sampling period in June, which coincided with the non-peak season, a total of 20 individuals (≈5 kg) were collected, with most specimens falling within the 200–300 g and > 300 g weight classes and total lengths predominantly between 60 and 80 cm, in accordance with the distributions summarized in Table 1.
Prigi Bay supports a diverse fishery, including large pelagic species such as Euthynnus affinis, Thunnus obesus, Thunnus alalunga, Makaira nigricans, Katsuwonus pelamis, Auxis rochei, and Auxis thazard. Small pelagic species such as Scomber australasicus, Sardinella lemuru, Sphyraena barracuda, Selar crumenophthalmus, Decapterus kurroides, and Rastrelliger kanagurta are also abundant, alongside demersal fishes including Trichiurus lepturus, Lutjanus johnii, Lates calcarifer, Leiognathus spp. , and Priacanthus spp.
The bay is relatively shallow (5–70 m) with muddy substrates and moderate sediment loads, conditions favorable for ribbonfish (10). Water quality in Prigi Bay remains within the standards of the Indonesian Ministry of Environment Decree No. 51/2004, with reported ranges of temperature (25.12–27.54 °C), salinity (33.75–34.54 ppt), and dissolved oxygen (6.73–8.87 ppm) (11). Variability in salinity and oxygen levels is influenced by depth, land-based inputs, and seasonal conditions. Environmental fluctuations such as reduced salinity, oxygen depletion, or pH decline can negatively affect marine organisms by disrupting respiration, growth, and survival (12). Poor water quality has further been associated with higher disease susceptibility in marine fish populations (13).
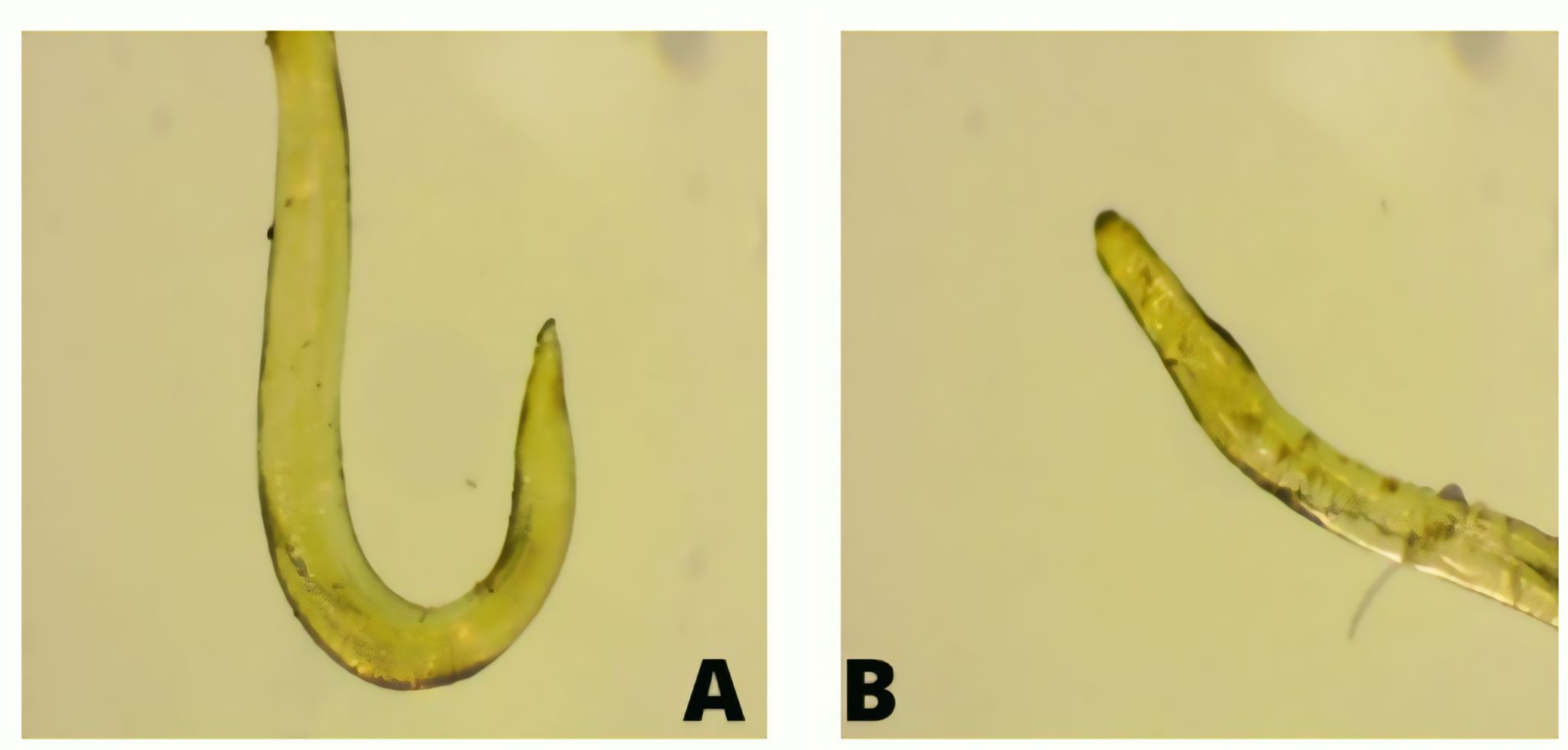
Endoparasites
A total of 56 ribbonfish were examined, consisting of 36 individuals from Paciran (Lamongan) and 20 individuals from Tamperan (Pacitan). The prevalence of endoparasite infection was 19.4% (7/36) in Paciran and 35.0% (7/20) in Tamperan. Infection intensity remained relatively low in both locations, with 1.3 individuals per infected fish in Paciran and 1.9 individuals per infected fish in Tamperan. Two nematode species, Anisakis sp. and Camallanus sp. , were detected in the gastrointestinal tract and muscle tissues. The results indicate that parasitic infection was more frequent in ribbonfish landed at Tamperan, although the overall parasitic burden was low.
Endoparasites are internal parasites commonly found in the gastrointestinal tract and visceral organs of fish, including the stomach, intestine, liver, and swim bladder. In ribbonfish, infections are often associated with food intake, which facilitates parasite transmission. Major groups of fish endoparasites include protozoa, digeneans, acanthocephalans, nematodes, and parasitic crustaceans (14). These infections can cause significant economic losses due to mortality, reduced growth, impaired condition, and increased susceptibility to secondary infections by fungi, bacteria, or viruses.
Clinical signs in infected fish generally include metabolic disorders, organ damage (particularly in the stomach and intestine), impaired growth, and increased mortality. Species frequently reported from ribbonfish include Eustrongylides sp. and Anisakis sp. in the stomach, and Acanthocephalus sp., Camallanus sp., Ascaris sp., and Trichuris trichiura in the intestine (15) Other nematodes, such as Capillaria sp. and acanthocephalans like Rhadinorhynchus sp. have also been documented (16).
Parasitic infections in fish are determined by the interplay of three major factors: the host, the parasite, and the environment. A well-functioning immune system enables fish to resist parasitic invasions, whereas poor environmental conditions can exacerbate infection risk. High organic matter, oxygen imbalance, and interspecific competition favor parasite persistence. Furthermore, inadequate water quality management and unhygienic feed, such as the use of untreated small fish as natural feed, have been reported to increase infection prevalence (16). Examples of ribbonfish specimens collected for parasitological examination are shown in Figure 1A.

The presence of parasites in fish is frequently associated with suboptimal water quality and other environmental stressors, which may reduce host condition and increase susceptibility to infection. Infected individuals often exhibit impaired growth and reduced body condition as parasites utilize nutrients from the gastrointestinal tract and other internal organs. However, in the present study, the relationship between host body weight and parasite burden was not formally tested using correlation analysis, and any apparent association should therefore be interpreted as suggestive rather than causative. Parasitological examination was performed using both external and internal inspections. External assessment included microscopic observation of fins, gills, and skin, while internal organs were examined following necropsy. Dissection was conducted using a scalpel from the abdominal cavity toward the pectoral fin, dorsal fin, and head. The stomach, intestine, liver, kidney, and muscle were removed, rinsed with physiological saline, and examined under a microscope. Gastrointestinal contents were emptied into Petri dishes to facilitate parasite identification.
Ribbonfish, being carnivorous, often prey on small fish and crustaceans, which serve as vectors for parasite transmission. In specimens collected from Prigi Bay, Trenggalek, small fish and crustaceans were found in the digestive tract, suggesting higher exposure to parasite sources. In contrast, specimens from Bawean waters, Gresik, exhibited smaller body weights and lacked prey remains in their intestines. This dietary difference may influence parasite prevalence and intensity. An example of prey items retrieved from the digestive tract is shown in Figure 1B.
The study identified two nematode endoparasites, Anisakis sp. and Camallanus sp. , in ribbonfish, localized in the intestine, stomach, and muscle tissues. Figure 2 presents the combined results of parasite abundance and the number of infected hosts across both landing sites. A total of 7 out of 36 fish from TPI Paciran Lamongan (19.4%) and 7 out of 20 fish from TPI Tamperan Pacitan (35.0%) were infected, yielding 14 infected individuals from 56 specimens examined. The higher prevalence and intensity observed in ribbonfish landed at Tamperan may be related to differences in feeding ecology and habitat use at Prigi Bay, where specimens more frequently contained small fish and crustaceans in their digestive tracts that can act as intermediate or paratenic hosts. This pattern suggests a potential link between trophic position and parasite exposure; however, because no formal statistical modeling of these relationships was conducted, the association remains indicative and should not be interpreted as demonstrating a direct causal effect of body size on infection risk.
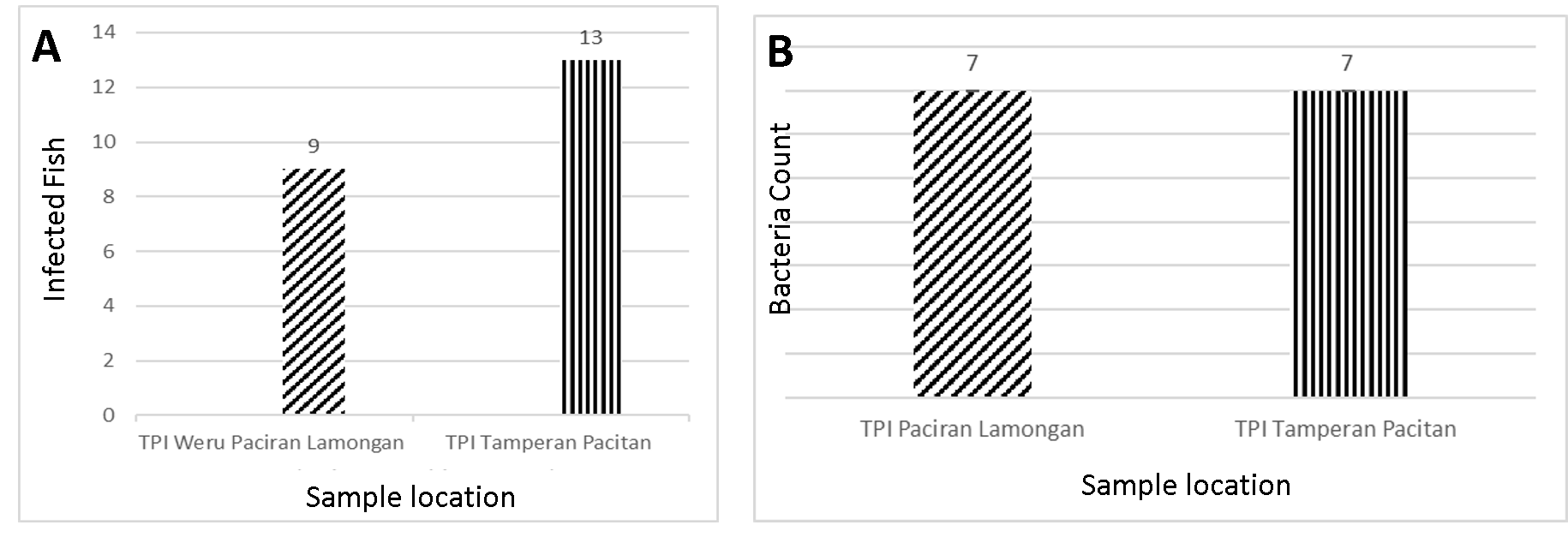
Anisakis sp.
One of the endoparasites identified in ribbonfish was Anisakis sp. The morphological features of this nematode are illustrated in Figure 3, showing both the anterior and posterior body regions.
Anisakis sp. is a nematode parasite characterized by an elongated cylindrical body, transparent in color, with a tapered posterior end. This parasite exhibits sexual dimorphism, with females possessing paired oviducts, ovaries, and a uterus, while males contain a single reproductive tract with testes, vas deferens, spicules, and a cloaca (17). As a zoonotic parasite, Anisakis sp. is capable of infecting marine mammals, fish, and occasionally humans.
In ribbonfish, Anisakis sp. penetrates the gastrointestinal tract and can migrate to other internal organs such as the stomach, intestines, and liver, where nutrient availability supports its survival. Heavy infections may reduce the host's body weight and overall condition. The life cycle begins when eggs are excreted in the feces of marine mammals, hatching into first-stage larvae (L1) in the water. After successive molts, free-swimming second-stage larvae (L2) are ingested by crustaceans, developing into third-stage larvae (L3). These larvae are then transmitted to fish as intermediate or paratenic hosts and finally reach adulthood in marine mammals. Larvae are inactivated at temperatures above 60 °C or below 0 °C (18).
Beyond ribbonfish, Anisakis sp. has also been reported in mackerel (Rastrelliger sp. ), tuna (Euthynnus sp. ), snapper (Lutjanus sp. ), and grouper, as well as cephalopods such as squid (19). This wide host range reflects the complex life cycle of Anisakis sp. and its ability to persist across multiple phic levels within marine ecosystems. Other zoonotic anisakid nematodes include Pseudoterranova, Hysterothylacium,Gnathostoma spinigerum, and Contracaecum, all of which are known to infect a wide range of marine and freshwater hosts and have been implicated in human parasitic infections. Transmission to humans typically occurs through the consumption of raw or undercooked seafood containing viable infective larvae. Once ingested, these larvae may invade the gastrointestinal tract, triggering clinical manifestations ranging from mild abdominal discomfort to severe gastrointestinal symptoms and allergic reactions. In certain cases, larval penetration and persistence may provoke inflammatory responses or hypersensitivity reactions, thereby exacerbating clinical severity. Consequently, the presence of zoonotic anisakid nematodes in seafood poses a significant risk for human anisakiasis and underscores the importance of proper seafood handling, processing, and cooking practices to reduce the risk of infection (18). The transmission mechanism of Anisakis sp. within the food web is illustrated in Figure 4.
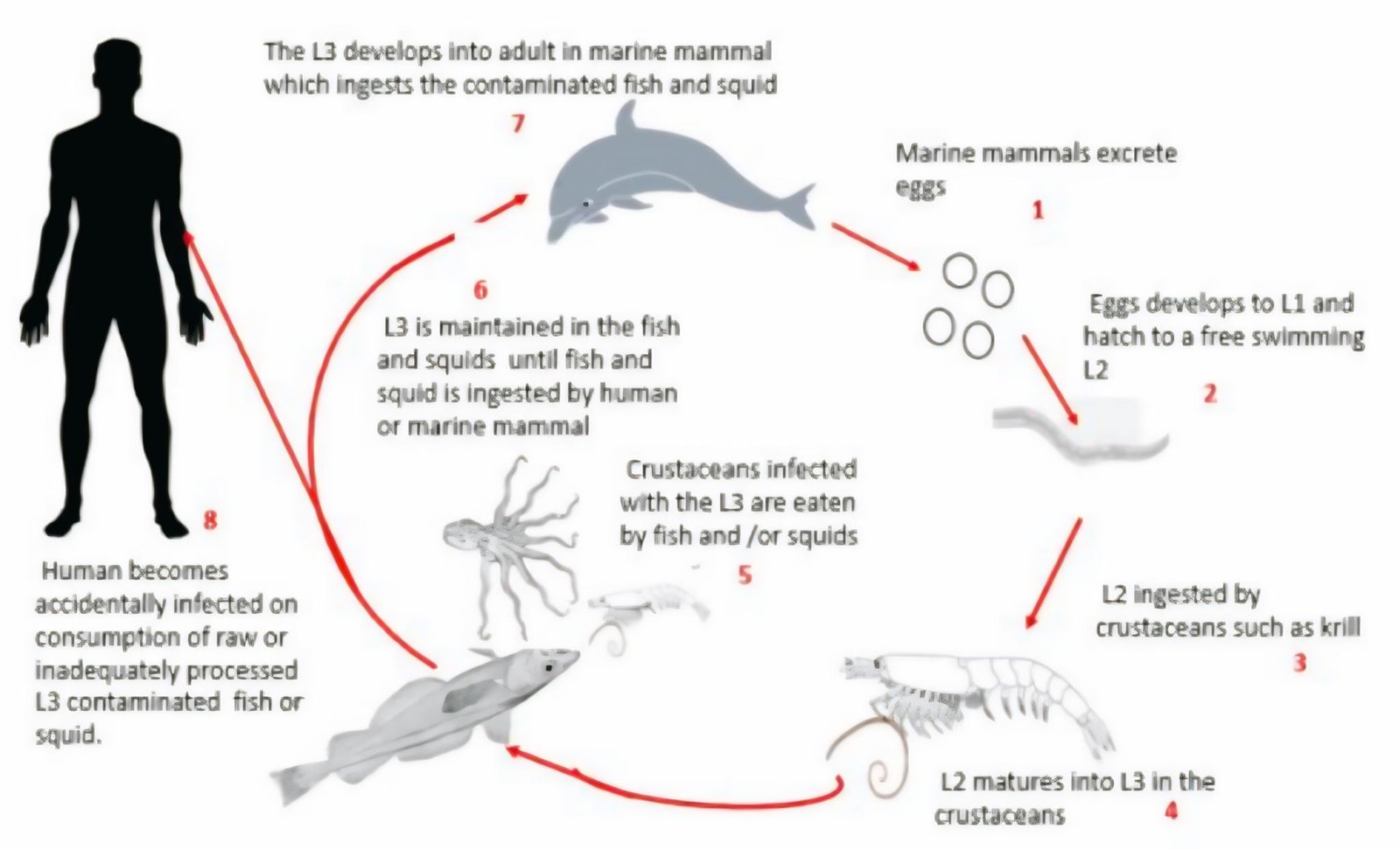
The life cycle of Anisakis sp. begins when marine mammals release eggs through feces, which hatch into first-stage larvae (L1) in seawater. These larvae develop into free-swimming L2 and are ingested by small fish or crustaceans, where they transform into infective third-stage larvae (L3). Larger fish and cephalopods become paratenic hosts by preying on these organisms, allowing the larvae to accumulate within their tissues. Humans and marine mammals are infected when consuming raw or undercooked seafood containing L3 larvae.
Infected fish may exhibit reduced body weight, gastrointestinal swelling, and impaired nutrient absorption. In humans, anisakiasis manifests with nausea, diarrhea, abdominal pain, allergic reactions, and in severe cases, eosinophilic granuloma or intestinal perforation (20).
Human cases are frequently reported in countries with high raw seafood consumption, such as Japan and parts of Europe. For instance, consumption of raw squid (sashimi, sushi) or inadequately processed anchovies (Engraulis encrasicolus) has been linked to outbreaks of anisakiasis, particularly caused by Anisakis pegreffii (18, 19).
Camallanus sp.
The second endoparasite identified in ribbonfish was Camallanus sp. , with its morphology presented in Figure 5.
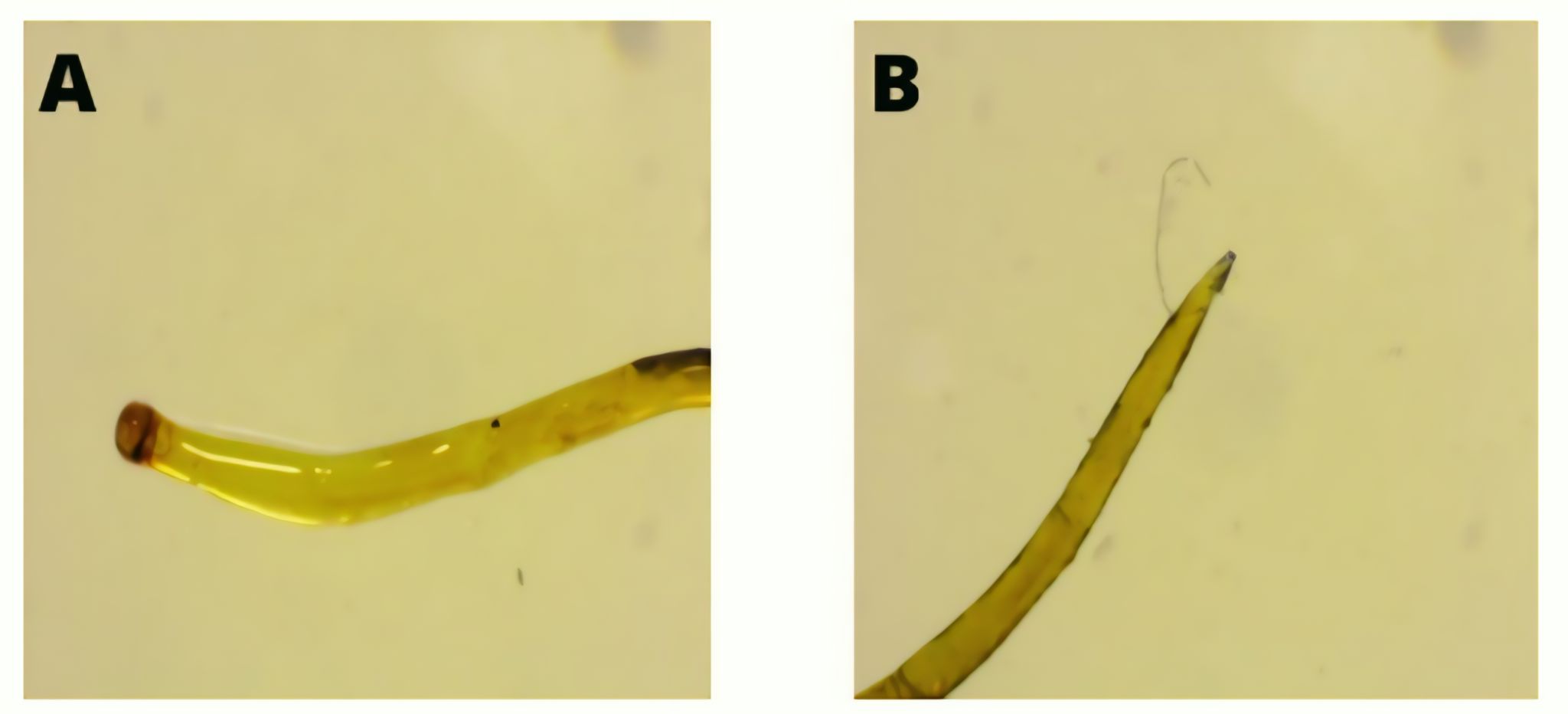
Camallanus sp. infects the intestine, pylorus, and caecum of fish, where it attaches to the mucosal lining and utilizes host nutrients for growth and reproduction. Taxonomically, it belongs to the phylum Nemathelminthes, class Nematoda, order Camallanoidea, family Camallaninae, and genus Camallaninae . The parasite is approximately 10 mm in length and is characterized by a cylindrical body covered by a cuticle extending from the anterior to the posterior region (mucron). The anterior end is rounded, whereas the posterior end is tapered, features that are commonly used as key diagnostic characters in morphological identification (21). The life cycle of Camallanus sp. is illustrated in Figure 6.
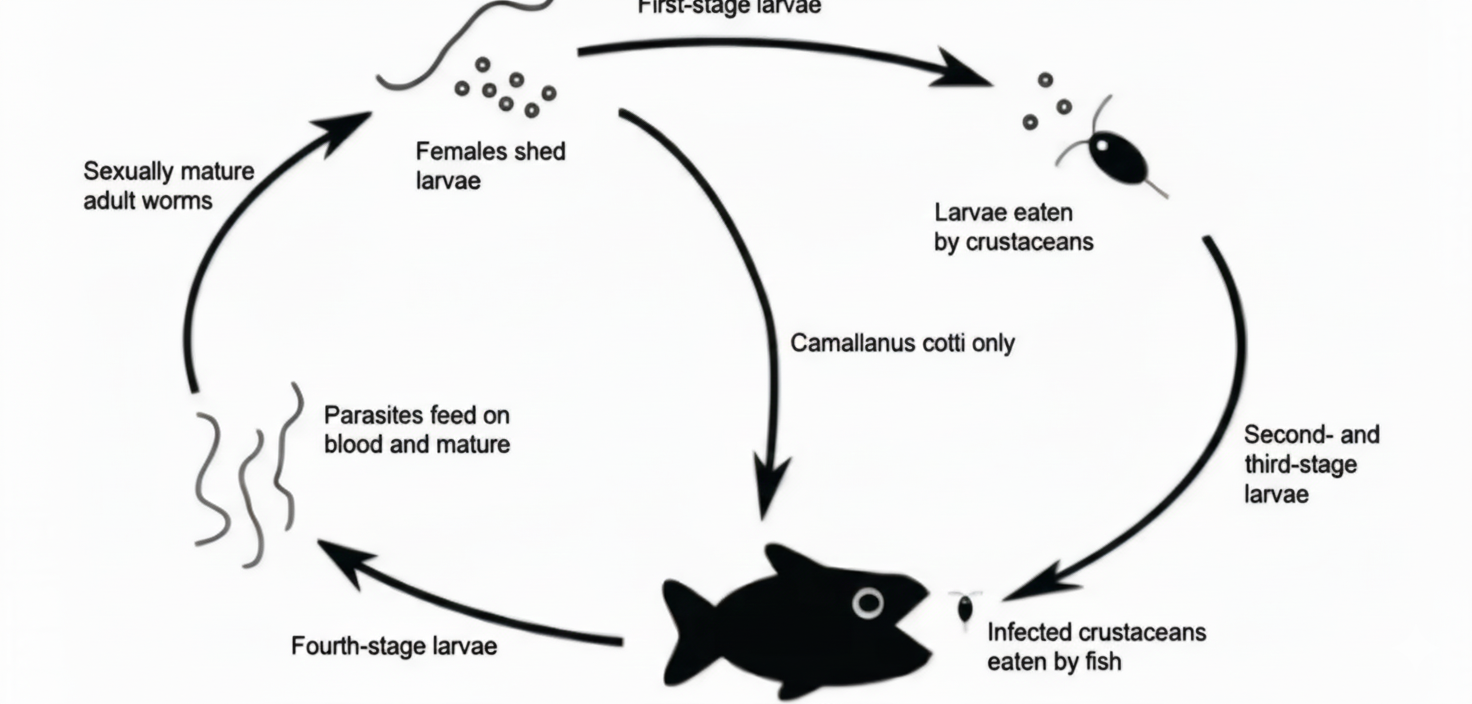
The life cycle begins with adult females releasing larvae into the host’s intestine. The larvae are expelled into the water, where they develop into the second-stage larvae (L2). These are ingested by crustaceans, serving as intermediate hosts, and further develop into infective third-stage larvae (L3). Once consumed by fish, the larvae mature into adults in the intestine. The larvae are capable of surviving in water for up to 12 days (22).
Although Camallanus sp. is not zoonotic like Anisakis sp. , it still causes significant harm to fish health. Clinical signs include intestinal and anal hemorrhage due to mucosal damage, anemia, deformities, and, in severe cases, mortality (23). Worms may protrude from the anus, and heavy infestations can permanently damage intestinal tissues (24).
Prevalence and Intensity of Parasites
The prevalence of endoparasite infection in ribbonfish and infection intensity is presented in Figure 7.
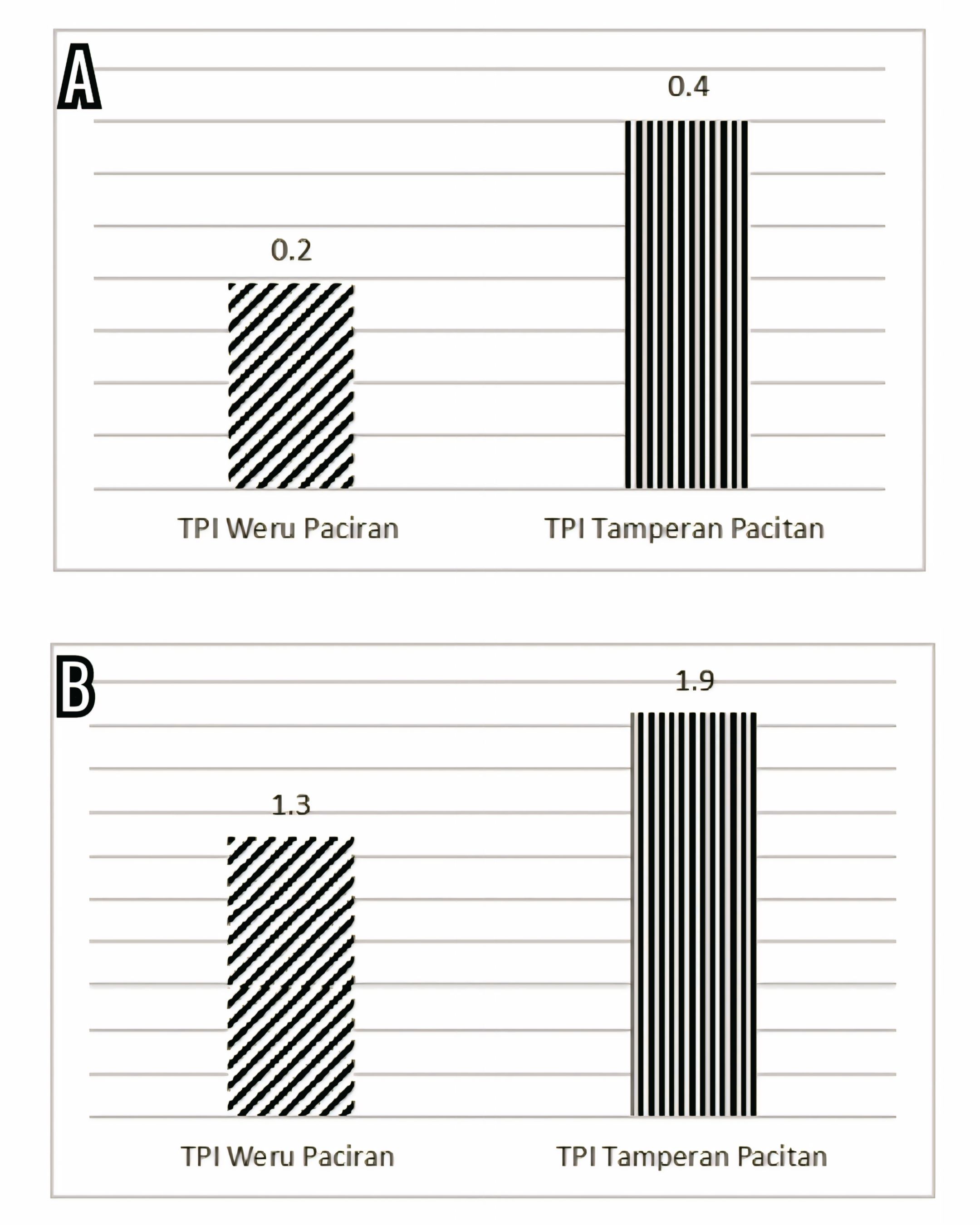
The prevalence of infection was 19.4% in ribbonfish landed at TPI Paciran (Lamongan) and 35.0% in those landed at TPI Tamperan (Pacitan). Infection intensity was 1.3 ind/individual in Paciran and 1.9 ind/individual in Tamperan, both categorized as low-level infections. Chi-square and Mann–Whitney U tests did not detect statistically significant differences in either prevalence or infection intensity between the two landing sites (p > 0.05), and therefore the results are mainly interpreted on the basis of descriptive patterns.
From Paciran, a total of 36 ribbonfish were examined, with 7 individuals infected by Anisakis sp. (body weight: 40–100 g; length: 40–50 cm). From Tamperan, 20 individuals were sampled, with 7 fish infected by Anisakis sp. and Camallanus sp. (body weight: 100–400 g; length: 60–80 cm).
Parasite distribution is influenced by environmental conditions, host size, age, sex, and other ecological factors (24). The higher prevalence and intensity in Tamperan samples may be associated with the larger body size of ribbonfish from Prigi Bay, which frequently contained small fish and crustaceans in their digestive tracts, serving as intermediate hosts.
Although infection levels in this study were relatively low compared to some other reports, they remain ecologically relevant. Prevalence and intensity values were lower compared to ribbonfish sampled from Cilacap, where infection reached 53.33% (25). Parasite infections are known to reduce appetite, growth, and metabolism, which may ultimately result in mortality (Nofyan, 2015) (26). However, host body weight alone is not a determining factor, as environmental conditions also play a crucial role in shaping infection dynamics (25).
Conclusion
This study confirmed the occurrence of two nematode endoparasites, Anisakis sp. and Camallanus sp. , infecting ribbonfish (Trichiurus lepturus) from Paciran and Tamperan landing sites. Although prevalence (19.4–35.0%) and infection intensity (1.3–1.9 ind/individual) were classified as low, the detection of zoonotic Anisakis sp. highlights the potential public health relevance of endoparasites in commercially important marine fish. Interpretation of the findings should consider several limitations, including the small sample size, the use of purposive rather than random sampling, the absence of molecular confirmation for parasite identification, and the lack of seasonal replication, which may have affected the generalizability of infection patterns. Future studies are recommended to incorporate longitudinal sampling across fishing seasons, molecular-level species validation, and expanded sampling coverage to better assess spatial and temporal variations in parasite transmission dynamics and evaluate associated risks for fisheries and human consumers.
Declarations
Conflict of Interest
The author declare no conflicting interest.
Data Availability
All data generated or analyzed during this study are included in this published article.
Ethics Statement
Ethical approval was not required for this study.
Funding Information
The author declare that no financial support was received for the research, authorship, and/or publication of this article.
References
- Indarjo A, Salim G, Amir F, Supriadi, Soejarwo PA, Nugraeni CD, et al. Growth characteristics of layur fish Lepturacanthus savala in Juata Waters, Tarakan, Indonesia. Ilmu Kelautan. 2020;25(3):127-134.
- Balai Penelitian dan Pengembangan Pengendalian Penyakit Bersumber Binatang Tanah Bumbu. Inventarization of parasite nematodes in plant, animal, and human. Media Litbang Kesehatan. 2017;13(3):195-207.
- Wulan Sari PD, Sri Subekti, Monica YD. Distribution of Anisakidae family–infected marine fish in Indonesia. Bioeduscience. 2021;5(3):188-195.
- Taghavimotlagh SA, Ghodrati Shojaei M, Vahabnezhad A. Life history traits of ribbonfish Trichiurus lepturus (Linnaeus, 1758) in the Persian Gulf and Oman Sea. Iranian Journal of Fisheries Sciences. 2021;20(2):298-312.
- Pardede M, Mahasri G, Ulkhaq MF. Intensity and prevalence of endoparasite helminths in little tuna (Euthynnus affinis) at Muncar and Panarukan Fish Auction Place, East Java, Indonesia. Iranian Journal of Aquatic Animal Health. 2020;6(2):49-59.
- Suman A, Irianto HE, Satria F, Amri DK. Penelitian Perikanan Laut. 2020.
- Yaskun M, Sugiarto E. Potensi hasil perikanan laut terhadap kesejahteraan para nelayan dan masyarakat di Kabupaten Lamongan. Jurnal Studi Manajemen dan Bisnis. 2017;4(1):257-264.
- Prihatiningsih P, Tirtadanu T, Taufik M, Rosmiati R, Nurdin E, Panggabean AS, et al. Unveiling the status of Trichiurus lepturus stocks in the southern Java waters, Indonesia: A biological and length-based assessment approach. Acta Ichthyologica et Piscatoria. 2025;55(1):31-43.
- Setyanto A, Rahman MA, Fatmasari Y, Irawan VB, Isdianto A, Asadi MA, et al. Species composition and length–weight frequency distribution of lobsters (Panulirus spp.) landed on the Puger Coast, Jember Regency, East Java. Berkala Sainstek. 2025;13(2):96-108.
- Fauziyah F, Akbarimansyah R, Ningsih EN, Agustriani F, Supriyadi F, Febrianti AAP. Pemetaan topografi dasar laut dan jenis sedimen di perairan Banyuasin, Sumatera Selatan. Jurnal Ilmu dan Teknologi Kelautan Tropis. 2023;15(2):251-264.
- Sidabutar EA. Distribusi suhu, salinitas, dan oksigen terlarut terhadap kedalaman di perairan Teluk Prigi Kabupaten Trenggalek. Journal of Fisheries and Marine Research. 2019;3(1):46-52.
- Ghazali M, Widoretno W, Arumingtyas EL, Retnaningdyah C. Macroepiphyte biodiversity on Kappaphycus alvarezii surface and its interaction with the environment in cultivation centers on Lombok Island, Indonesia. Biodiversitas. 2022;23(12):6284-6292.
- LeChevallier MW, Prosser T, Stevens M. Opportunistic pathogens in drinking water distribution systems: A review. Microorganisms. 2024;12(5):916.
- Simarani K. Endoparasites inventory and prevalence of Scomberomorus commerson in Sumatera Province, Indonesia. Egyptian Journal of Aquatic Biology and Fisheries. 2024;28(4):1373-1383.
- França LF, Knoff M, da Fonseca MCG, Gomes DC, Ferreira MS, Felizardo NN, et al. Lecithochirium monticellii (Digenetic trematode) parasites of Trichiurus lepturus from the State of Rio de Janeiro, Brazil, with notes on taxonomy. Anais da Academia Brasileira de Ciências. 2020;92(2):1-7.
- Linayati L, Mardiana TY, Adimahsyaf NF, Yahya MZ, Yuniarti T. Derajat infeksi dan prevalensi ektoparasit pada ikan mas (Cyprinus carpio) di Kabupaten Batang. Jurnal Perikanan Unram. 2024;14(2):781-788.
- Bello E, Palomba M, Webb SC, Paoletti M, Cipriani P, Nascetti G, et al. Multilocus genotyping reveals interspecific hybridization of Anisakis pegreffii and A. berlandi in southern Pacific Ocean waters. Infection, Genetics and Evolution. 2021;92:1.
- Aibinu IE, Smooker PM, Lopata AL. Anisakis nematodes in fish and shellfish – from infection to allergies. International Journal for Parasitology: Parasites and Wildlife. 2019;9:384-393.
- Cipriani P, Giulietti L, Shayo SD, Storesund JE, Bao M, Palomba M, et al. Anisakid nematodes in Trichiurus lepturus and Saurida undosquamis from the South-West Indian Ocean: Genetic evidence for sister species within Anisakis typica (s.l.). Food and Waterborne Parasitology. 2022;28:e00177.
- Cong W, Elsheikha HM. Biology, epidemiology, clinical features, diagnosis, and treatment of selected fish-borne parasitic zoonoses. Yale Journal of Biology and Medicine. 2021;94(2):297.
- Abdel-Gaber R, Maher S, Abdel-Gaber R. Morphological re-description of Camallanus polypteri (Nematoda: Camallanidae) infecting African sharptooth catfish Clarias gariepinus. Archives of Razi Institute. 2021;76(2):303-309.
- Hamid A, Lamondo D, Aydalina RV. Prevalensi, intensitas, dan dominansi parasit pada ikan manggabai (Glossogobius giuris) di Danau Limboto Provinsi Gorontalo. Bioscientist: Jurnal Ilmiah Biologi. 2024;12(1):508-516.
- Kamoura A, Ismail I, Mohamady N, El Alem M. Effect of Camallanus spp. infestation on hematological, immunobiochemical and histological changes in catfish (Clarias gariepinus) with treatment trials. Animal Health Research Journal. 2020;8(3).
- Veronika E, Elvince R, Wulandari L. Parasite infection, prevalence, intensity and dominance in climbing perch (Anabas testudineus) in Sebangau River, Central Kalimantan, Indonesia. Aquaculture, Aquarium, Conservation & Legislation (AACL Bioflux). 2024;17(2):843-852.
- Wu S, Wang G, Gao D, Xi B, Yao W, Liu M. Occurrence of Camallanus cotti in diverse fish species from Danjiangkou Reservoir in central China. Parasitology Research. 2007;101(2):467-471.
- Ihsan B, Sitinjak RS. Prevalence of ectoparasites in tilapia (Oreochromis niloticus) in Tarakan. Journal of Medical and Veterinary Sciences. 2023;6(2):185-190