RESEARCH ARTICLE
Microplastic Distribution and Dynamics Between Riparian Vegetation and Alaknanda River in the Himalayan Region
Aquatic Life Sciences|Vol. 3, Issue 2, pp. 52-61 (2026)
CC BY 4.0-2026 Authors
Views
Downloads
Shares
Received
Jan 27, 2026Revised
Mar 13, 2026Accepted
Jun 24, 2026Published
Jul 10, 2026
Abstract
Microplastic (MP) contamination in freshwater ecosystems is an emerging environmental concern, yet interactions between MPs and riparian macrophytes remain poorly understood, particularly in Himalayan river systems. This study provides a baseline assessment of MP surface entanglement across riparian macrophyte species, along with associated surface water and sediment, from River Alaknanda, Srinagar, Uttarakhand, India, sampled in June 2023. A total of 30 macrophyte, surface water, and sediment samples were collected and processed using extraction protocols integrating washing, oxidative digestion, density separation, stereomicroscopic analysis, and polymer characterization through ATR-FTIR spectroscopy. MPs were extensively detected on macrophyte surfaces, with species-specific normalized burdens ranging from 16.47 ± 2.59 to 273.86 ± 26.21 MPs/g dry weight. Kruskal-Wallis analysis revealed significant interspecific variation in normalized MP abundance (H = 25.73, p = 0.0023), with Chenopodium ambrosioides exhibiting significantly higher MP burdens than Conyza sp. and Equisetum sp. No MPs within the analytical detection range of 100 µm-5 mm were detected in digested plant tissues. Fibres were the dominant MP type in macrophytes and water, whereas fragments predominated in sediments, with white particles being most abundant across all matrices. Polymer analysis identified polyester, polyethylene, polypropylene, polyterephthalate, and polyethylene-vinyl acetate as dominant polymers. Strong compositional correlations among macrophytes, water, and sediment indicated similarities in MP characteristics across environmental matrices. These findings suggest riparian macrophytes as complementary matrices for freshwater MP monitoring. Despite methodological and spatial limitations, this study establishes a preliminary framework for incorporating riparian vegetation into freshwater MP monitoring and highlights the need for further investigations.
Introduction
Freshwater ecosystems are increasingly recognized as significant sinks and transport pathways for microplastics (MPs), which are now widely reported in rivers, lakes, groundwater, sediments, and aquatic biota worldwide (1-4). Rivers, in particular, play a crucial role in the transport, transformation, and deposition of MPs, linking terrestrial plastic sources with downstream aquatic environments (3, 5). Within riverine ecosystems, MPs have been documented in surface water, sediments, and diverse organisms including macroinvertebrates, fish, and aquatic vegetation (5-7). Despite growing attention to freshwater MP contamination, interactions between MPs and riparian or aquatic macrophytes remain comparatively understudied (8-10).
Riparian macrophytes are important structural components of freshwater ecosystems, contributing to habitat complexity, sediment stabilization, and nutrient cycling (11-13). They occupy the interface between terrestrial and aquatic environments and are periodically exposed to suspended particles transported by river flow. Because their surfaces remain in direct contact with water, sediments, and floating debris, they may accumulate MPs and therefore represent a potentially useful environmental matrix for evaluating MP distribution within freshwater ecosystems. Previous studies have documented MP associations with aquatic vegetation, although such investigations remain limited in freshwater river systems.
Previous studies investigating MP-macrophyte associations have largely focused on marine ecosystems, particularly seagrasses and macroalgae, where surface adhesion of MPs has been documented (8-10). However, freshwater systems, especially Himalayan riverine ecosystems, remain poorly studied in this regard. Current knowledge gaps include the extent of MP surface entanglement on freshwater riparian vegetation, potential internal accumulation within plant tissues, and the relationship between MPs associated with macrophytes and those present in surrounding water and sediment compartments.
The River Alaknanda, a major headstream of the River Ganga, is ecologically and socioeconomically significant and increasingly subjected to anthropogenic pressures including urbanization, tourism, domestic waste discharge, and hydropower development (5, 7). These pressures may contribute to MP contamination, particularly in riparian zones where vegetation interacts directly with riverine flow and suspended pollutants (5, 17). Understanding MP distribution across riparian vegetation and adjacent environmental matrices in such systems is essential for establishing baseline ecological assessments.
Therefore, this study aimed to: (i) assess the occurrence and extent of MP surface entanglement across riparian macrophyte species in River Alaknanda; (ii) evaluate potential internal accumulation of MPs within plant tissues; (iii) characterize MP abundance, shape, color, and polymer composition in macrophytes, water, and sediment; and (iv) examine relationships between MPs detected in macrophytes and surrounding environmental matrices. We hypothesized that MP abundance would vary among macrophyte species and that MP characteristics observed on macrophytes would broadly reflect patterns present in adjacent environmental matrices. Because quantitative morphometric characterization of plant surfaces was beyond the scope of this study, mechanisms responsible for interspecific differences were not directly evaluated.
Material and Methods
Study Area
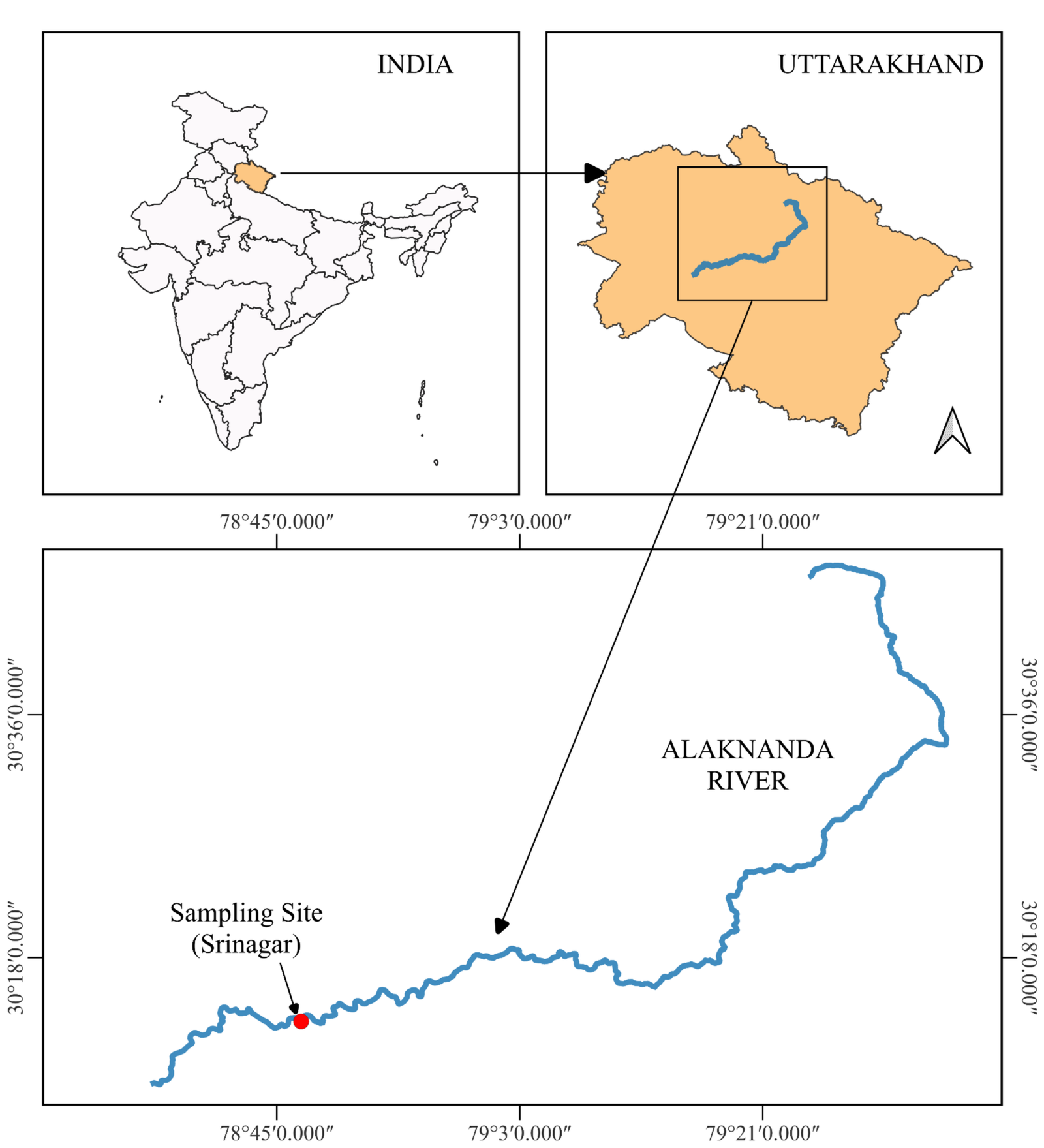
A stretch of the River Alaknanda at Srinagar (Garhwal), Uttarakhand, India was selected for this study. The Alaknanda River is an integral part of the Ganga River system; it meets River Bhagirathi at Devprayag to form the River Ganga. It is a perennial river regulated by the Satopanth glacier in the Himalayas and seasonal rainwater. The Alaknanda River flourishes major towns like Badrinath, Joshimath, Chamoli, Karnaprayag, Rudraprayag, and Srinagar. It flows more than 150 km within the deep mountain gorges and hilly terrain, confluencing with some major and minor tributaries like Ghrit Ganga, Saraswati, Dhauli Ganga, Nandakini, Birahi Ganga, Pindar and Mandakini before reaching Srinagar. Srinagar is a highly populated town and is considered an educational hub of the hilly region. The river water is used for hydropower projects, religious and drinking purposes. The selected study region near Srinagar represents an ecologically and anthropogenically relevant riparian zone influenced by urban settlement, tourism, domestic sewage discharge, transportation activities, and religious practices, all of which may contribute to MP inputs.
The river stretch in this region was selected due to the presence of well-established riparian macrophyte communities, direct anthropogenic pressures, and accessibility for simultaneous sampling of vegetation, water, and sediment. These factors make the site suitable for assessing baseline MP distribution across multiple environmental compartments. The riparian zone along the Alaknanda River hosts perennial macrophyte communities, enduring seasonal flooding for approximately four months each year. Beyond its hydrological and environmental significance, the river is also a habitat for a rich variety of flora and fauna. The surrounding region is known for its biodiversity, with diverse aquatic and terrestrial species contributing to the ecosystem. Notable fauna includes Himalayan Snow Trout, Mahseer species, and stone loach, while the riverbanks are adorned with an array of plant life. This confluence of diverse species within the Alaknanda River ecosystem adds an additional layer of complexity and ecological importance to the study area, highlighting the intricate interdependence between macrophytes and the broader river ecosystem.
Sampling
For this study, sampling was conducted in the month of June 2023 near Govt Polytechnic College, Srinagar, Uttarakhand, India (30° 13' 17.0364'' N and 78° 46' 48.1620'' E) from two riparian patches along River Alaknanda (Figure 1). Ten species of macrophytes (underwater) were identified and collected along the river bank with 3 replicates of each species (Table 1). The plant was cut using a scissor and packed into aluminium foil and transported to the Aquatic Ecology laboratory. Along with plant samples, water and sediment samples were also taken from the same region. For surface water samples, 500 L of water was filtered immediately using 100-micron and 5 mm sieves and the extracted sample was collected through backwashing of sieves in vials (18). For sediment sample, three 500 g (wet sediment) replicates were collected from the same area as the water and macrophyte in airtight glass jar and carried back to the laboratory (19). Identification of these plants was conducted using an online website, called as, ‘Plant of world online’(20).
| Macrophyte Species | Number | Wet Weight (g) (Mean ± SD) | Dry Weight (g) (Mean ± SD) |
|---|---|---|---|
| Amaranthus sp. | 3 | 4.7 ± 0.4 | 0.9 ± 0.3 |
| Chenopodium ambrosioides | 3 | 9.8 ± 1.6 | 1.5 ± 0.2 |
| Conyza sp. | 3 | 120.8 ± 20 | 18 ± 4 |
| Panicum sp. | 3 | 54 ± 6 | 11 ± 1.8 |
| Equisetum sp. | 3 | 12.8 ± 2.4 | 3.6 ± 0.4 |
| Poa sp. | 3 | 10.1 ± 1.8 | 2.3 ± 0.3 |
| Solanum nigrum | 3 | 45.4 ± 9 | 8.5 ± 1.4 |
| Tamarix aphylla | 3 | 24.3 ± 3.2 | 6.6 ± 0.5 |
| Xanthium strumarium | 3 | 38.3 ± 2.3 | 5.9 ± 0.25 |
| Bidens Pilosa | 3 | 13.5 ± 4.8 | 8.1 ± 4.9 |
MP Extraction
For further extraction of MPs from the samples, the water sample was treated with hydrogen peroxide (30%) at 70 °C (21, 22), density separated using ZnCl2 and filtered using gridded cellulose nitrate filter paper with pore size 0.8 microns (Himedia) (Figure 2). For sediment samples, wet sediment was dried in a hot air oven at 75 °C (23), and 100 g sediment was taken for further analysis i. e., density separation and filtration.
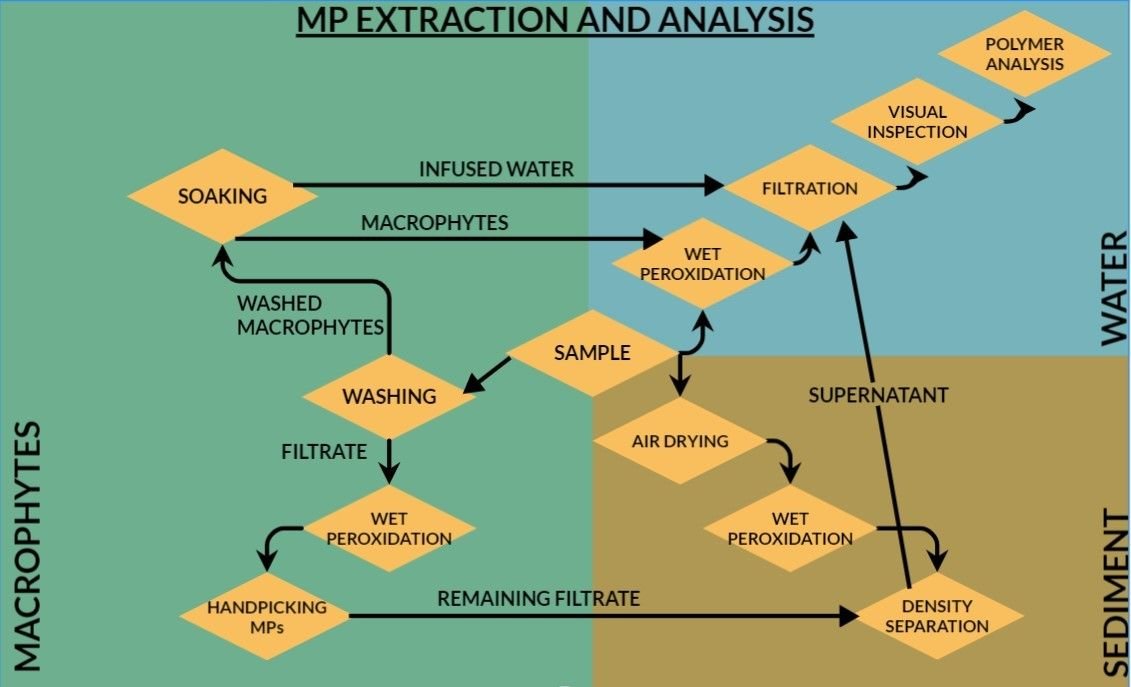
Because no standardized protocol currently exists for simultaneous assessment of surface-entangled and internally accumulated MPs in freshwater riparian macrophytes, a customized methodological workflow was developed by integrating established freshwater MP extraction procedures with modified plant tissue processing approaches. Oxidative digestion using H₂O₂ was selected based on its demonstrated efficiency in removing plant-derived organic matter while maintaining acceptable polymer recovery in plant and algal matrices (24, 25).
Freshly collected macrophyte samples were first processed to isolate surface-associated MPs. Each sample was carefully washed three times using double-distilled water over a 100 µm stainless-steel sieve to remove externally entangled particles. The retained wash filtrate was collected through backwashing and preserved for subsequent MP extraction. To further recover loosely adhered particles, washed macrophytes were immersed in double-distilled water for one hour, after which the infused water was filtered and retained for analysis.
Following surface processing, wet biomass of each macrophyte sample was recorded. Samples were then oven-dried at 75 °C until constant weight to determine dry biomass. Dried plant tissues were subsequently subjected to oxidative digestion using 30% hydrogen peroxide at 70 °C for 6-7 days to degrade organic material and assess potential internalized MPs. After digestion, samples were filtered for microscopic examination. This two-stage workflow enabled separate evaluation of surface-entangled MPs and potential internal tissue accumulation.
MP Analysis
The recovered particles were analysed for their physical shape, and color under a stereomicroscope at 40× magnification for preliminary identification and enumeration. According to visible morphological characteristics particles were classified into fragments, fibres, films, clusters and foam, and further classified on the basis of color. Larger suspected particles were manually isolated where feasible, and hot needle testing was applied in uncertain cases to distinguish synthetic particles from non-plastic materials. From the total MPs detected across all sample types, a random subset comprising approximately 5% of particles from each matrix (water, sediment, and macrophytes) was selected for polymer validation, resulting in 455 analyzed particles. Of these, 432 particles were confirmed as synthetic plastic polymers through spectroscopic analysis, corresponding to a validation efficiency of approximately 95%. Based on this confirmation rate, an estimated 5% analytical error margin was considered in MP count interpretation to improve overall data reliability.
Polymer identification was performed using a Shimadzu IR spectrometer (Model A224159) equipped with an IRSpirit™ unit and a QATR-S single-reflection ATR attachment fitted with a diamond crystal. Semi-transparent and transparent particles were analyzed in transmission mode, whereas opaque particles were examined in ATR mode after mounting on gold-coated glass microscope slides. Spectral measurements were acquired over the range of 4000-400 cm⁻¹, with a 3-second collection time, 10 co-scans per sample, and a constant spectral resolution of approximately 0.9 cm⁻¹. Aperture dimensions were adjusted between 50 × 50 µm² and 150 × 150 µm² depending on particle size. Each obtained spectrum was compared against polymer reference libraries for polymer classification. This integrated morphological and chemical characterization framework enhanced confidence in MP identification while recognizing that complete spectroscopic validation of all recovered particles was beyond the scope of this exploratory baseline study.
Data Analysis
Descriptive statistics, including means, standard deviations, ranges were used to summarize MP occurrence among sampled macrophyte species and environmental compartments. To improve interspecific comparability, MP abundance in macrophytes was normalized relative to both wet and dry biomass (MPs/g wet weight and MPs/g dry weight). Dry weight-normalized values were used as the primary metric for species-level comparisons due to reduced variability associated with plant water content.
Because sample sizes were limited and data did not meet assumptions for robust parametric testing, non-parametric statistical analyses were employed. Differences in normalized MP abundance among macrophyte species were evaluated using the Kruskal-Wallis H test. Where significant overall differences were detected, Dunn’s post hoc pairwise comparisons were performed to identify specific interspecific differences.
Correlation analyses were conducted to compare MP shape and color distributions among macrophytes, water, and sediment, thereby assessing compositional associations among environmental matrices. Pearson correlation coefficients (r) were used to estimate the strength of these compositional associations. Data tabulation, normalization, graphical visualization, and statistical analyses were conducted using Microsoft Excel and supplementary statistical software tools.
Quality Control
To minimize contamination during field and laboratory procedures, glassware, filtration units, and metal instruments were thoroughly rinsed with distilled water prior to use. Macrophyte samples were wrapped in aluminum foil immediately after collection, and direct contact with plastic materials was minimized wherever feasible. Cotton fabrics, latex gloves, and glassware were utilized during both field and laboratory work to reduce synthetic contamination risk. All instruments involved in the extraction process underwent thorough cleaning with distilled water and subsequent drying before analysis. Field blanks and airborne contamination controls were not incorporated into this exploratory design. However, procedural blanks were included and corresponding corrections were applied during data interpretation. Consequently, potential background contamination, particularly for fibre-type particles cannot be entirely excluded.
Results
MPs Occurrence in Macrophytes
In this study, ten plant species, namely, Amaranthus sp., C. ambrosioides, Conyza sp., Panicum sp., Equisetum sp., Poa sp., S. nigrum, T. aphylla, X. strumarium, B. pilosa (Figure 3) were assessed for the presence of MPs. A total of 8, 812 MP particles were recovered from the surface washings of ten riparian macrophyte species sampled from River Alaknanda, indicating widespread surface entanglement of MPs across all examined vegetation. Fibres constituted the dominant particle shape (81.8%), followed by fragments (18.0%), while clusters represented a minor fraction (0.2%). An image showing different MPs in washed sample is mentioned as Supplementary Figure 1. Observed clusters comprised intricate entanglements, each composed of hundreds of multi-colored fibres (Supplementary Figure 2). White-colored particles were the most prevalent across all species, although multiple additional colors including black, red, yellow, blue, green, pink, and purple were also observed. On an individual basis, MP entanglement ranged from 149 to 747 particles per plant.

Dry Weight Normalization
Normalized MP abundance differed markedly among species, ranging from 16.47 ± 2.59 MPs/g dry weight in Equisetum sp. to 273.86 ± 26.21 MPs/g dry weight in C. ambrosioides (Figure 4). Amaranthus sp. also exhibited high normalized MP burden (230.26 ± 76.40 MPs/g dry weight), followed by Poa sp. (99.62 ± 54.21 MPs/g dry weight) and X. strumarium (96.61 ± 27.11 MPs/g dry weight). Lower burdens were recorded in Conyza sp. (17.40 ± 7.78 MPs/g dry weight) and Equisetum sp.
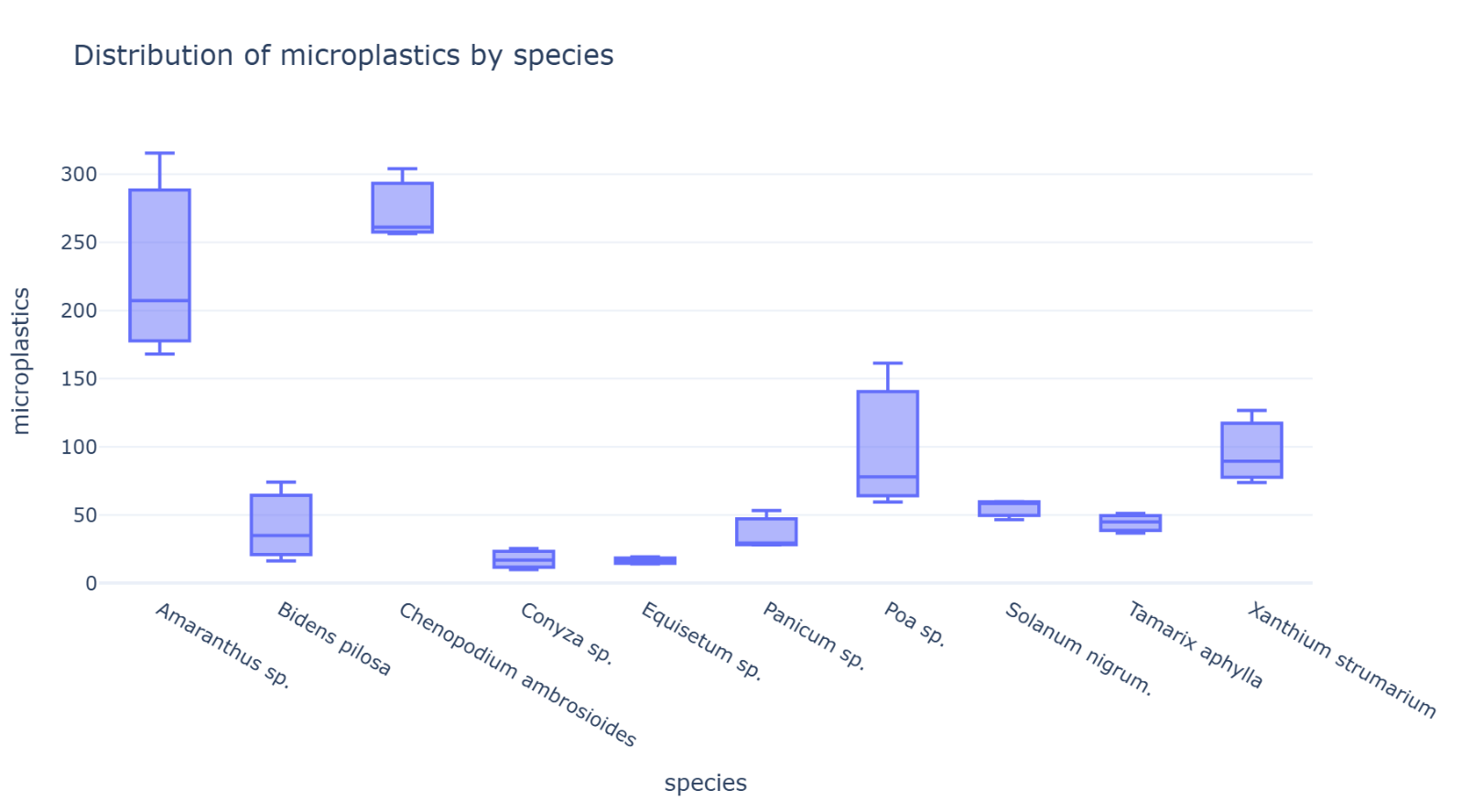
Kruskal-Wallis analysis revealed statistically significant interspecific differences in normalized MP abundance (H = 25.73, df = 9, p = 0.0023). Post hoc Dunn's test with Bonferroni correction revealed that C. ambrosioides exhibited significantly higher MP abundance than Conyza sp. (Z = 3.29, adjusted p = 0.0447) and Equisetum sp. (Z = 3.39, adjusted p = 0.0320). No other pairwise comparisons remained significant following Bonferroni correction (Supplemental Figure 1).
Wet Weight Normalization
Patterns observed on a wet weight basis were consistent with dry weight findings. C. ambrosioides (43.51 ± 6.26 MPs/g wet weight) and Amaranthus sp. (40.75 ± 5.10 MPs/g wet weight) retained the highest normalized MP burdens, while Conyza sp. (2.55 ± 1.06 MPs/g wet weight) exhibited the lowest.
Internalized MPs in Macrophyte Tissues
No MPs were detected within digested macrophyte tissues within the analytical size range evaluated (100 μm-5 mm). Therefore, while larger MPs were not observed within plant tissues, the occurrence of smaller MPs or nanoplastics cannot be excluded.
MPs in Water and Sediment
A total of 231 MPs were detected in the water sample, categorized into fibres (60.2%), fragments (36.8%), film (2.2%), and foam (0.8%). Remarkably, white MPs were the most prevalent, constituting 45.4% of the observed MPs in the water sample- amounting to 105 MPs. Apart from white MPs, various colors were also identified, including 29 blue, 27 black, 25 green, 17 red, 15 pink, and 13 yellow particles (Figure 5).
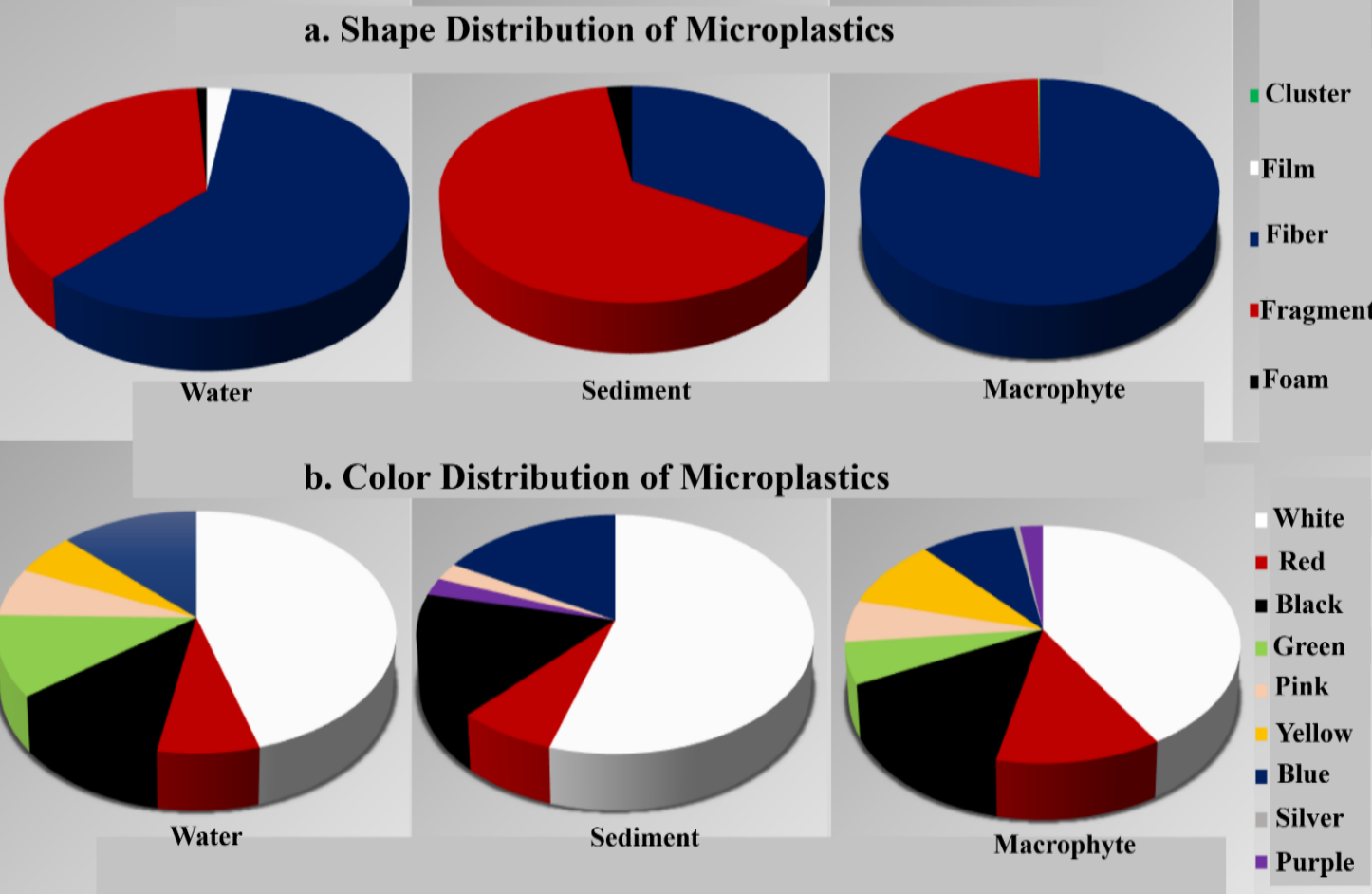
In the sediment sample, a total of 42 MP particles were identified. Within this count, 33.3% were classified as fibres, 64.3% as fragments, and 2.4% as foam. White MP emerged as the predominant color, amounting to 23 MPs i. e. , 54.8% of overall observation in the sediment sample. Apart from white MPs, a spectrum of colored MPs was also observed. Specifically, 7 particles each of blue and black, 3 of red, and 1 each of pink and purple were discerned.
Correlation between MPs in Macrophytes, Water and Sediment
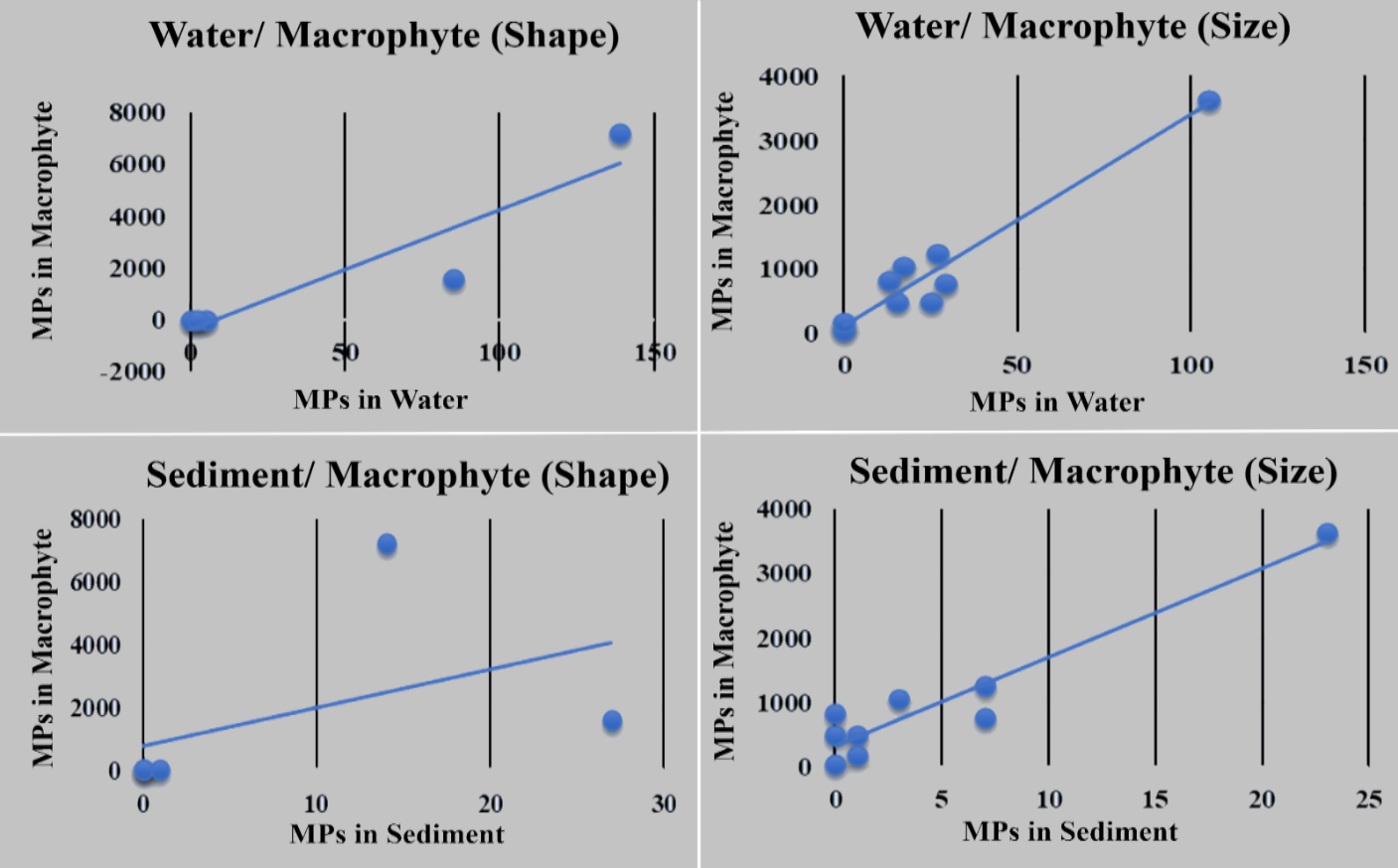
Pearson correlation analysis demonstrated strong positive associations in MP color composition between macrophytes and water (r = 0.967, p < 0.001) and between macrophytes and sediment (r = 0.953, p < 0.001) (Supplemental Figure 2). Shape composition also showed a significant positive correlation between macrophytes and water (r = 0.917, p = 0.028). In contrast, the correlation between macrophytes and sediment based on particle shape was moderate and not statistically significant (r = 0.443, p = 0.455) (Figure 6). These findings indicate compositional similarities among environmental matrices. However, correlation analyses alone cannot establish pollution sources, transport pathways, or mechanistic relationships among matrices.
Polymer Analysis
Six types of polymers were found in the polymer analysis of MPs, i. e. , Polyester, High-density Polyethylene, Low-density Polyethylene, Polypropylene, Polyterephthalate and Polyethylene-Vinyl acetate. However, a few particles were identified as Kolophonium-rosin. A figure depicting different type of polymers is given as Supplementary Figure 3.
Discussion
This study provides baseline evidence of widespread MP surface entanglement across ten riparian macrophyte species in a Himalayan freshwater ecosystem, highlighting that MPs are not restricted solely to water and sediment compartments but are also extensively associated with riverbank vegetation. The detection of MPs on all examined macrophyte species suggests that riparian vegetation may represent an important, yet comparatively understudied, environmental interface for MP accumulation within freshwater systems. While previous investigations have primarily focused on MP occurrence in water, sediments, and aquatic fauna, comparatively limited attention has been directed toward macrophyte-associated MPs, particularly in riverine freshwater habitats, a few studies conducted in marine ecosystems highlighted such interactions (9, 10).
Biomass-normalized analyses revealed substantial interspecific variation in MP burden, with C. ambrosioides and Amaranthus sp. exhibiting significantly greater MP loads compared to several other species. These findings demonstrate substantial interspecific variation in MP abundance among riparian macrophytes. Although substantial interspecific variation was observed, the present study did not quantify morphological variables such as leaf area, trichome density, branching architecture, or surface roughness. Consequently, the observed variation cannot be directly attributed to specific structural traits. Future studies integrating quantitative morphometric measurements with MP abundance data would help clarify the mechanisms underlying species-specific accumulation patterns. However, the processes governing this accumulation require further investigation.
Other than plant morphology, surface adhesion of MPs may also be influenced by biofilms, polysaccharide layers, hydrodynamic conditions, and environmental exposure factors, which have been identified in previous aquatic studies (14-16). Seasonal submergence of riparian vegetation during monsoonal flow may increase opportunities for MP entanglement, particularly in river systems such as the Alaknanda where fluctuating hydrological conditions may influence plastic transport and retention. During monsoon period the study site remains submerged, allowing the entanglement of plastic waste to the macrophytes which is clearly notable during post-monsoon period (Supplementary Figure 4). However, these potential mechanisms were not directly measured in the present study and should therefore be considered as plausible explanatory pathways rather than confirmed processes.
No MPs were detected within digested macrophyte tissues under the analytical conditions employed, suggesting that within the detectable size range, internal accumulation of larger MPs was absent. This finding should be interpreted cautiously, as analytical limitations, including detection thresholds and potential nanoplastic presence, may constrain definitive conclusions regarding complete absence of internalized plastic particles. While larger MPs may be excluded by plant structural barriers, finer-scale studies employing advanced detection methods are needed to evaluate nanoplastic uptake potential more comprehensively.
In our investigation, MPs were distinguished based on their shape and color. Predominantly, white fibres were identified in samples collected from washing macrophyte and water samples, while white fragments dominated in sediment samples. Despite the absence of industrial pollution sources, Srinagar is experiencing significant MP contamination, primarily from domestic sewage. The abundance of MP fibres in the River Alaknanda is attributed to factors such as high domestic effluent, open waste dumping, atmospheric deposition, wastewater discharge, recreational activities, runoff, and tourism, given the river's proximity to the Badrinath National Highway. Correspondingly, recent research on the Ganga River, from Rishikesh, Uttarakhand to Farakka, West Bengal, revealed the prevalence of white fibres, particularly those composed of nylon and polyester (17). The dominance of white color may stem from various factors, including the types of plastics used in the region, particle decoloration through photodegradation, pigment loss, and their residential duration in the river, along with wet peroxidation during laboratory procedures. Nevertheless, the MP pollution depends on the study area, pollution levels, and environmental factors. Providing a singular trend regarding the availability of MPs according to their color and shape in a particular aquatic body is challenging, as it is influenced by numerous associated factors.
In this investigation, positive correlations were observed between the shape and color characteristics of MPs associated with macrophytes and those present in adjacent water and sediment matrices. These relationships indicate compositional similarities among environmental compartments. Similar MP characteristics may arise from multiple environmental inputs and processes. Correlation alone cannot establish common pollution sources or direct transport pathways and would require additional approaches such as polymer fingerprinting, source-tracking analyses, or hydrological investigations.
The polymer analysis identified polyester, polyethylene, polypropylene, polyterephthalate, and polyethylene-vinyl acetate among dominant polymer groups. These polymers are commonly associated with consumer products, packaging materials, textiles, and household waste, suggesting diffuse anthropogenic activities as likely contributors to MP contamination. However, precise source attribution was beyond the scope of the present study, and polymer occurrence should be interpreted as reflective of multiple potential inputs rather than definitive pollution origins.
Several methodological limitations, including single-season sampling, limited spatial coverage, small replication size, absence of comprehensive contamination controls, and partial FTIR validation, constrain the broader generalization of these findings. Additionally, direct quantitative comparisons among macrophytes, water, and sediment remain limited by differing sample units and matrix characteristics. Therefore, this study should be considered an exploratory baseline assessment rather than definitive ecological quantification. Nevertheless, the results demonstrate that riparian macrophytes can retain substantial quantities of MPs on their surfaces, with species-specific differences in MP abundance.
Conclusion
This study provides preliminary baseline evidence that riparian macrophytes in the River Alaknanda can act as surfaces for MP entanglement. MPs were extensively detected on the external surfaces of all examined macrophyte species, with substantial interspecific variation in normalized MP burden, indicating that species-specific factors may influence MP accumulation patterns. Biomass-normalized analyses and statistical testing demonstrated significant differences among species, with certain macrophytes exhibiting disproportionately greater MP burdens. No MPs were detected within digested plant tissues under the analytical conditions employed, suggesting that within the detectable size range, MP occurrence was primarily restricted to external surfaces rather than internal accumulation. However, methodological limitations, including particle-size detection thresholds, warrant cautious interpretation of this finding.
The predominance of fibres and shared compositional patterns among macrophytes, water, and sediment indicate that riparian vegetation contains MP assemblages broadly similar to those observed in surrounding environmental matrices. However, broader validation across species, seasons, and environmental conditions is required before such applications can be established. Polymer analyses further revealed the presence of commonly used anthropogenic polymers, suggesting diffuse consumer-related sources. While the study is constrained by limited temporal and spatial replication, small sample size, and exploratory methodological design, it establishes an important preliminary framework for understanding MP associations with freshwater riparian vegetation. Future research should incorporate broader seasonal sampling, enhanced contamination controls, detailed morphological analyses, and advanced particle detection approaches to better resolve the mechanisms and ecological significance of MP-macrophyte interactions.
Declarations
Conflict of Interest
The authors declare no conflicting interest.
Data Availability
Data supporting the findings of this study are available from the corresponding author upon reasonable request.
Ethics Statement
Ethical approval was not required for this study.
Funding Information
The authors declare that no financial support was received for the research, authorship, and/or publication of this article.
Supplemental Material
<b> Supplemental Figures 1–4</b> are available in a single file at the following <a class="cursor-pointer" href="https://etflin.com/file/document/20260619200117_651904_aac25b31.docx">link</a>.
References
- Amrutha K, Warrier AK. The first report on the source-to-sink characterization of microplastic pollution from a riverine environment in tropical India. Science of The Total Environment. 2020;739:140377. doi: https://doi.org/10.1016/j.scitotenv.2020.140377
- Bhatt V, Badola N, Semwal D, Singh Chauhan J. Status of Microplastic Pollution in the Freshwater Ecosystems. Singapore: Springer Nature Singapore; 2023. doi: https://doi.org/10.1007/978-981-99-5018-8_7
- Han M, Niu X, Tang M, Zhang BT, Wang G, Yue W, et al. Distribution of microplastics in surface water of the lower Yellow River near estuary. Science of The Total Environment. 2020;707:135601. doi: https://doi.org/10.1016/j.scitotenv.2019.135601
- Panno SV, Kelly WR, Scott J, Zheng W, McNeish RE, Holm N, et al. Microplastic Contamination in Karst Groundwater Systems. Groundwater. 2019;57(2):189-196. doi: https://doi.org/10.1111/gwat.12862
- Chauhan JS, Semwal D, Nainwal M, Badola N, Thapliyal P. Investigation of microplastic pollution in river Alaknanda stretch of Uttarakhand. Environ Dev Sustain. 2021;23(11):16819-16833. doi: https://doi.org/10.1007/s10668-021-01388-y
- Badola N, Sobhan F, Chauhan JS. Microplastics in the River Ganga and its fishes: Study of a Himalayan River. Science of The Total Environment. 2023;901:165924. doi: https://doi.org/10.1016/j.scitotenv.2023.165924
- Bhatt V, Badola N, Chauhan JS. Microplastic in fishes: the first report from a Himalayan River — Alaknanda. Environ Sci Pollut Res. 2023;31(1):1637-1643. doi: https://doi.org/10.1007/s11356-023-30889-8
- Seng N, Lai S, Fong J, Saleh MF, Cheng C, Cheok ZY, et al. Early evidence of microplastics on seagrass and macroalgae. Marine and Freshwater Research. 2020;71(8):922-928. doi: https://doi.org/10.1071/mf19177
- Goss H, Jaskiel J, Rotjan R. Thalassia testudinum as a potential vector for incorporating microplastics into benthic marine food webs. Marine Pollution Bulletin. 2018;135:1085-1089. doi: https://doi.org/10.1016/j.marpolbul.2018.08.024
- Datu SS, Supriadi S, Tahir A. Microplastic in Cymodocea rotundata Seagrass Blades. Ijeab. 2019;4(6):1758-1761. doi: https://doi.org/10.22161/ijeab.46.21
- Christie H, Norderhaug K, Fredriksen S. Macrophytes as habitat for fauna. Mar. Ecol. Prog. Ser. 2009;396:221-233. doi: https://doi.org/10.3354/meps08351
- Dhote S. Role of Macrophytes in improving water quality of an aquatic eco-system. Journal of Applied Sciences and Environmental Management. 2007;11(4):133-5.
- Ladislas S, El-Mufleh A, Gérente C, Chazarenc F, Andrès Y, Béchet B. Potential of Aquatic Macrophytes as Bioindicators of Heavy Metal Pollution in Urban Stormwater Runoff. Water Air Soil Pollut. 2011;223(2):877-888. doi: https://doi.org/10.1007/s11270-011-0909-3
- Oberbeckmann S, Loeder MG, Gerdts G, Osborn AM. Spatial and seasonal variation in diversity and structure of microbial biofilms on marine plastics in Northern European waters. FEMS Microbiology Ecology. 2014;90(2):478-492. doi: https://doi.org/10.1111/1574-6941.12409
- Gutow L, Eckerlebe A, Giménez L, Saborowski R. Experimental Evaluation of Seaweeds as a Vector for Microplastics into Marine Food Webs. Environ. Sci. Technol. 2015;50(2):915-923. doi: https://doi.org/10.1021/acs.est.5b02431
- Rummel CD, Jahnke A, Gorokhova E, Kühnel D, Schmitt-Jansen M. Impacts of Biofilm Formation on the Fate and Potential Effects of Microplastic in the Aquatic Environment. Environ. Sci. Technol. Lett. 2017;4(7):258-267. doi: https://doi.org/10.1021/acs.estlett.7b00164
- Rajan K, Khudsar FA, Kumar R. Urbanization and population resources affect microplastic concentration in surface water of the River Ganga. Journal of Hazardous Materials Advances. 2023;11:100342. doi: https://doi.org/10.1016/j.hazadv.2023.100342
- Hidalgo-Ruz V, Gutow L, Thompson RC, Thiel M. Microplastics in the Marine Environment: A Review of the Methods Used for Identification and Quantification. Environ. Sci. Technol. 2012;46(6):3060-3075. doi: https://doi.org/10.1021/es2031505
- Blair RM, Waldron S, Phoenix VR, Gauchotte-Lindsay C. Microscopy and elemental analysis characterisation of microplastics in sediment of a freshwater urban river in Scotland, UK. Environ Sci Pollut Res. 2019;26(12):12491-12504. doi: https://doi.org/10.1007/s11356-019-04678-1
- Royal Botanic Gardens K. Plants of the World Online 2026 [Available from: https://powo.science.kew.org/.
- Li J, Liu H, Paul Chen J. Microplastics in freshwater systems: A review on occurrence, environmental effects, and methods for microplastics detection. Water Research. 2018;137:362-374. doi: https://doi.org/10.1016/j.watres.2017.12.056
- Al-Azzawi MSM, Kefer S, Weißer J, Reichel J, Schwaller C, Glas K, et al. Validation of Sample Preparation Methods for Microplastic Analysis in Wastewater Matrices—Reproducibility and Standardization. Water. 2020;12(9):2445. doi: https://doi.org/10.3390/w12092445
- Zhang D, Fraser MA, Huang W, Ge C, Wang Y, Zhang C, et al. Microplastic pollution in water, sediment, and specific tissues of crayfish (Procambarus clarkii) within two different breeding modes in Jianli, Hubei province, China. Environmental Pollution. 2021;272:115939. doi: https://doi.org/10.1016/j.envpol.2020.115939
- Prihandari R, Karnpanit W, Kittibunchakul S, Kemsawasd V. Development of Optimal Digesting Conditions for Microplastic Analysis in Dried Seaweed Gracilaria fisheri. Foods. 2021;10(9):2118. doi: https://doi.org/10.3390/foods10092118
- Pfeiffer F, Fischer EK. Various Digestion Protocols Within Microplastic Sample Processing—Evaluating the Resistance of Different Synthetic Polymers and the Efficiency of Biogenic Organic Matter Destruction. Front. Environ. Sci. 2020;8. doi: https://doi.org/10.3389/fenvs.2020.572424