RESEARCH ARTICLE
Phytochemical Profiling with Biological Validation Reveals Therapeutic Effect of Vincetoxicum capparidifolium
Sciences of Phytochemistry|Vol. 5, Issue 2, pp. 232-248 (2026)
Views
Downloads
Shares
Received
Apr 16, 2026Revised
Jun 14, 2026Accepted
Jun 26, 2026Published
Jul 11, 2026
Abstract
The present research discusses the phytochemical composition, anti-inflammatory, antidiabetic, and antiproliferative efficacy of Vincetoxicum capparidifolium leaf aqueous extract. Phytochemical characterization was performed using FTIR and LC-MS analysis. Network pharmacology was employed to identify potential molecular targets of tylophorine associated with liver associated disorders, followed by molecular docking studies. Anti-inflammatory activity and antidiabetic potential was evaluated in vitro. Cytotoxic outcomes were determined using MTT assay on HepG2 cells, along with AO/EtBr staining and DNA fragmentation analysis. FTIR investigation disclosed the occurrence of various functional groups, incorporating hydroxyl, amine, aromatic, and heteroatom-containing moieties. LC-MS profiling categorized a total of 28 compounds belonging to alkaloids, flavonoids, phenols, and fatty acid derivatives. Network pharmacology analysis identified 94 intersecting targets of tylophorine with liver inflammation, diabetic liver disease, and end-stage liver disease, while molecular docking showed binding affinities of tylophorine with proteins, presenting the strongest interaction with 3HHM (-8.9 kcal mol-1). The extract produced concentration-dependent inhibition of protein denaturation (9.5-68.0%), proteolytic activity (10.3-71.7%), and erythrocyte lysis (10.6-70.2%) although its activity was lower than the reference drug, aspirin. The extract also displayed inhibition of α-amylase and α-glucosidase, with greater potency against α-glucosidase (IC50=99.6 µg mL-1). The cytotoxic activity evaluated using MTT assay supported reduction in HepG2 cell viability (IC50=168 µg mL-1). AO/EtBr staining revealed increased apoptotic features, including membrane damage and nuclear condensation, while DNA fragmentation analysis verified apoptosis-mediated cell death. Overall, V. capparidifolium exhibits notable in vitro anti-inflammatory, antidiabetic, and cytotoxic potential, highlighting its potential as a source of bioactive compounds for further pharmacological investigations.
Keywords:
Introduction
A major aspect in the onset of cancer is chronic inflammation, predominantly for hepatocellular carcinoma, where persisting inflammatory signaling triggers angiogenesis, tumor initiation, proliferation, and resistance to intervention (1, 2). Reactive oxygen species, chemokines, and cytokines are manifestations of pro-inflammatory mediators that are fundamental in establishing a microenvironment that promotes tumor growth and accelerates the development of disease (2, 3). While standard chemotherapy approaches have facilitated the clinical management of cancer, limitations such as systemic toxicity, acquired resistance, and patient response variability may have an impact on their long-term efficacy (4). These drawbacks highlight the need of creating multi-target treatment strategies that can safely modify important inflammatory and carcinogenic pathways (5, 6).
Natural products' unique structural variety and tendency to interact with several biochemical targets have attracted considerable research interest as possible therapeutic resources (7, 8). They could influence a variety of biological processes owing to their chemical complexity, frequently with lower toxicity and higher biocompatibility than synthetic drugs. It has been widely shown that secondary metabolites originating from plants, such as alkaloids, flavonoids, and phenolic chemicals, have anti-inflammatory and anticancer properties (9-11). These compounds act through multiple mechanisms, including the regulation of key signaling pathways, suppression of oxidative stress, inhibition of pro-inflammatory mediators, and induction of apoptosis in cancer cells (10, 11). Moreover, their ability to concurrently affect many sites renders them particularly advantageous in tackling intricate, multifactorial disorders like cancer (9). Because of their dual purpose, plant-derived metabolites are compelling choices for integrative therapy approaches meant to treat malignancies linked to inflammation.
Numerous indigenous plant species with great phytochemical variety may be found in the Western Ghats (12-15). Among these, members of the family Apocynaceae are especially notable for their abundance of phytocompounds, such as alkaloids, flavonoids, and terpenoids, which possess major pharmacological properties (16-19). Despite this, several endemic species within this family remain underexplored for their therapeutic potential. Vincetoxicum capparidifolium (Wight & Arn. ) Kuntze, an endemic species of the Western Ghats, holds ethnomedicinal importance, particularly in the traditional treatment of jaundice (20, 21). Although our recent study demonstrated the phytochemical richness and cytotoxic activity of the ethanolic leaf extract of V. capparidifolium, the phytochemical composition and biological properties of its aqueous extract remain unexplored (22). Since extraction solvent could influences metabolite recovery and biological activity, investigation of the aqueous extract may reveal distinct phytochemical constituents and pharmacological effects. There is also an apparent dearth of literature on the genus Vincetoxicum's in vitro anti-inflammatory and antidiabetic capacity. Therefore, the present study aimed to characterize the phytochemical profile of the aqueous leaf extract of V. capparidifolium using FTIR and LC-MS analyses and to evaluate its anti-inflammatory, antidiabetic, and cytotoxic activities through in vitro assays. In addition, in silico approaches were employed to explore potential molecular targets of the compound tylophorine associated with liver-related disorders and to provide preliminary mechanistic insights into the observed biological activities.
Experimental Section
Plant Collection and Extraction
Fresh leaves of V. capparidifolium were collected from Thaishola, Nilgiris district, Tamil Nadu, India on April
2025. The collected specimen was authenticated from Botanical Survey of India, Southern Circle, Coimbatore, India
(BSI/SRC/5/23/Tech-623). This authentication process serves as a critical quality control step to ensure taxonomic
precision. The specimens were washed and wiped systematically to remove dirt. The samples were shade dried,
processed, and kept in air-tight plastic packets. The powdered leaf material of V. capparidifolium was
extracted by cold maceration in distilled water (1: 10 w/v) for 72 h at room temperature with intermittent
shaking. This gentle method was specifically selected to prevent the thermal degradation of heat-sensitive
bioactive compounds. The extract was filtered and the filtrate was frozen and subsequently lyophilized to obtain
a dry aqueous extract. The extraction yield was calculated based on the weight of the lyophilized extract relative to
the initial dry plant material and was found to be 18.9 %. The lyophilized extract was stored at -20 ℃ until further
use (23).
FTIR Analysis
FTIR spectroscopy of the V. capparidifolium leaf aqueous extract was performed using a Shimadzu IR Affinity-1S FTIR spectrometer (Shimadzu Corporation, Japan) equipped with a deuterated triglycine sulfate detector. Spectra were recorded in the mid-infrared region of 400-4000 cm-1 at a spectral resolution of 4 cm-1. Baseline correction was applied to minimize spectral noise and the resulting absorption bands were analyzed and assigned by comparison with standard reference spectra to identify the functional groups present (24).
LC-MS QTOF Analysis
LC-MS QTOF analysis of the V. capparidifolium leaf aqueous extract was conducted using a dual AJS ESI Q-TOF system (G6550A) in positive ion mode, employing an AutoMS2 acquisition mode with a scan range of m/z 120-1200 for MS and medium isolation width of 4 amu for MS/MS. The binary pump delivered a gradient elution at 0.300 mL min-1 (pressure limit 1200 bar): 5% B (0.1% formic acid in water) initially, ramped to 95% B (acetonitrile) over 25-30 min, held until 35 min, with source parameters including sheath gas temperature at 350℃, gas flow at 13 L min-1, nebulizer at 35 psig, VCap at 3500 V, nozzle at 1000 V, fragmentor at 175 V, skimmer at 65 V, and octopole RF peak at 750 V. MS/MS parameters featured a relative threshold of 5% (absolute 0.010), precursor exclusion after 0.20 min and 1 spectrum, maximum 10 precursors per cycle targeting 25, 000 counts/spectrum, and ramped collision energy (slope 4, offset -2.6) for charges +1 to +3. Metabolites were putatively identified based on accurate mass, retention time, isotope patterns, and spectral database matching (25).
Network Pharmacology and Molecular Docking Approach
Genes allied with liver inflammation, diabetic liver disease, and end-stage liver disease were recovered from the GeneCards database (26). The complete disease-associated gene sets were used for subsequent analyses without applying an initial relevance score threshold. Tylophorine, pinpointed through LC-MS analysis of the V. capparidifolium aqueous extract, was chosen as the representative compound for its consistent detection in both +ve and -ve ionization modes. Potential targets of tylophorine were envisaged with SwissTargetPrediction database (27). The predicted targets of tylophorine were intersected with the disease-associated gene sets to identify common targets. The overlapping targets were imported into the STRING database to construct a protein-protein interaction network and enrichment analysis (28). The network was further analyzed in Cytoscape, where topological parameters including degree, closeness centrality, and radiality were calculated and the top 10 hub genes were identified (29). A composite scoring system was developed to prioritize candidate genes by integrating normalized network topological parameters (degree, closeness centrality, and radiality) with disease relevance scores using a weighted linear model adapted from established disease-gene prioritization frameworks that combine network topology and biological knowledge sources (30, 31). Disease relevance was assigned the highest weight (0.30) to emphasize genes with stronger documented associations to liver disorders, while degree and closeness centrality were each assigned weights of 0.25 because of their importance in network connectivity and information flow. Radiality was assigned a weight of 0.20 as a complementary measure of network influence (32, 33).
Genes were ranked based on the composite scores, and the top 3 candidates were selected for molecular docking analysis. Three proteins corresponding to the prioritized hub genes were retrieved from the RCSB Protein Data Bank. Molecular docking was performed using PyRx software and protein structures were prepared by Molegro Molecular Viewer. The ligand (tylophorine) was energy-minimized prior to docking and the best conformations were selected based on docking scores. The docked complexes were visualized using Discovery Studio to analyze binding interactions and conformational orientation of the ligand within the active site (34, 35).
In Vitro Anti-inflammatory Effect
The anti-inflammatory potential of the aqueous leaf extract of V. capparidifolium was evaluated using protein denaturation, anti-proteinase, and human red blood cell (HRBC) membrane stabilization assays. In the protein denaturation assay, the reaction mixture consisted of egg albumin (0.2 mL), phosphate-buffered saline (PBS, pH 6.4; 2.8 mL), and plant extract (0.6 mL) at concentrations of 10-250 µg mL-1 prepared in 0.2% DMSO. The mixtures were incubated at 37 °C for 10 min, heated at 70 °C for 20 min, cooled, and absorbance was recorded at 660 nm (36). The anti-proteinase activity was assessed using a reaction mixture containing trypsin (250 µL), 25 mM Tris-HCl buffer (pH 7.4; 1.0 mL), and plant extract (10-250 µg mL-1). Following pre-incubation at 37 °C for 5 min, casein (1.0 mL, 0.8% w/v) was added and incubation continued for 20 min. The reaction was terminated with 70% perchloric acid (2.0 mL), centrifuged, and absorbance of the supernatant was measured at 280 nm (37). HRBC membrane stabilization was evaluated using human blood collected in EDTA, centrifuged, washed with isotonic saline, and reconstituted as a 10% (v/v) suspension. Reaction mixtures (1 mL) containing HRBC suspension (100 µL) and plant extract (10-250 µg mL-1) were incubated at 54 °C for 30 min, centrifuged, and absorbance was measured at 560 nm (38). Aspirin served as the reference standard in all assays, appropriate controls were maintained, and experiments were performed in triplicate.
In Vitro Antidiabetic Capacity
The antidiabetic potential of the aqueous leaf extract of V. capparidifolium was evaluated through α-amylase and α-glucosidase inhibitory assays using acarbose as the reference standard. In the α-amylase inhibition assay, Starch Azure (2 mg) was suspended in 0.5 M Tris-HCl buffer (pH 6.9) containing 0.01 M CaCl2 (0.2 mL), boiled for 5 min, and preincubated at 37 °C for 5 min. The plant extract (0.2 mL; 10-250 µg mL-1) was added, followed by porcine pancreatic α-amylase (0.1 mL; 2 U mL-1). After incubation at 37 °C for 10 min, the reaction was terminated using 50% acetic acid (0.5 mL), centrifuged at 3000 rpm for 5 min (4 °C), and absorbance of the supernatant was recorded at 595 nm (39). The α-glucosidase inhibitory activity was determined in a 96-well plate containing phosphate buffer (50 µL, 100 mM, pH 6.8), α-glucosidase (10 µL; 1 U mL-1), and plant extract (20 µL; 10-250 µg mL-1). Following preincubation at 37 °C for 15 min, p-nitrophenyl-α-D-glucopyranoside (20 µL; 5 mM) was added as substrate and incubated for 20 min. The reaction was terminated with 0.1 M Na2CO3 (50 µL), and absorbance of the released p-nitrophenol was measured at 405 nm. All experiments were performed in triplicate with appropriate controls (39).
In Vitro Characterization of Cell Viability
Dulbecco's Modified Eagle Medium enriched with 10% fetal bovine serum (FBS), 100 µg mL-1 streptomycin, and 100 µg mL-1 penicillin was selected to cultivate the human liver cancer cell line (HepG2) provided by the National Centre for Cell Sciences (NCCS), Pune. The media was replaced every three days while the cells resided at 37 ℃ in an incubator with 5% CO2. HepG2 cells were seeded in 96-well plates at a density of 15,000 cells per well for MTT experiment. The cells were subsequently grown for 48 h at 37 ℃ with 5% CO2. Evaluations were conducted on V. capparidifolium leaf aqueous extract at 125, 250, 500, and 1000 µg mL-1 concentrations. Doxorubicin-treated cells were the positive control, and untreated cells were the negative control. Under an inverted microscope, morphological changes were noted over a 24-hour period. After that, 100 µL of new media containing 10 µL of MTT reagent (5 mg mL-1) was added to the culture medium, and it was cultured for four hour. A Multiskan SkyHigh Plate Reader was used to detect absorbance at 570 nm after the formazan crystals were dissolved in 100 µL of DMSO. Plotting a dose-response curve of log concentration against cell survival percentage allowed for the calculation of cell viability and the determination of the IC50 value (40).
AO/EtBr Staining
Approximately 5 µL of dye mixture containing acridine orange (AO) and ethidium bromide (EtBr) at a concentration of 100 mg mL-1 each was added to 9 mL of the cell suspension (1×105 cells/mL) on a clean microscopic coverslip. The mixture was incubated for 2-3 min, after which the cells were observed under a fluorescence microscope (Nikon Eclipse, Inc. , Japan) at 40× magnification using an excitation filter of 510-590 nm. The percentage of apoptotic cells was then calculated (41).
DNA Fragmentation
HepG2 cells were planted in 6-well plates at a density of 1×106 cells mL-1 in the DMEM with 10% fetal bovine serum (FBS), 100 µg mL-1 streptomycin, and 100 µg mL-1 penicillin in order to investigate internucleosomal DNA cleavage. After that, the cells were exposed to a fixed dose of plant extract, and as a positive control, 10 µg mL-1 of actinomycin D was utilized. Following the specified incubation periods, the cells were extracted, and agarose gel electrophoresis was used to measure DNA fragmentation (41).
Statistical Analysis
All experiments were performed in triplicate, and data are presented as mean ± standard deviation (SD). Statistical analyses were performed using Microsoft Excel. Differences among groups were evaluated by one-way analysis of variance (ANOVA) followed by Tukey's post hoc test. Differences were considered statistically significant at p< 0.05.
Results and Discussion
Functional Groups Identified from V. capparidifolium Extract
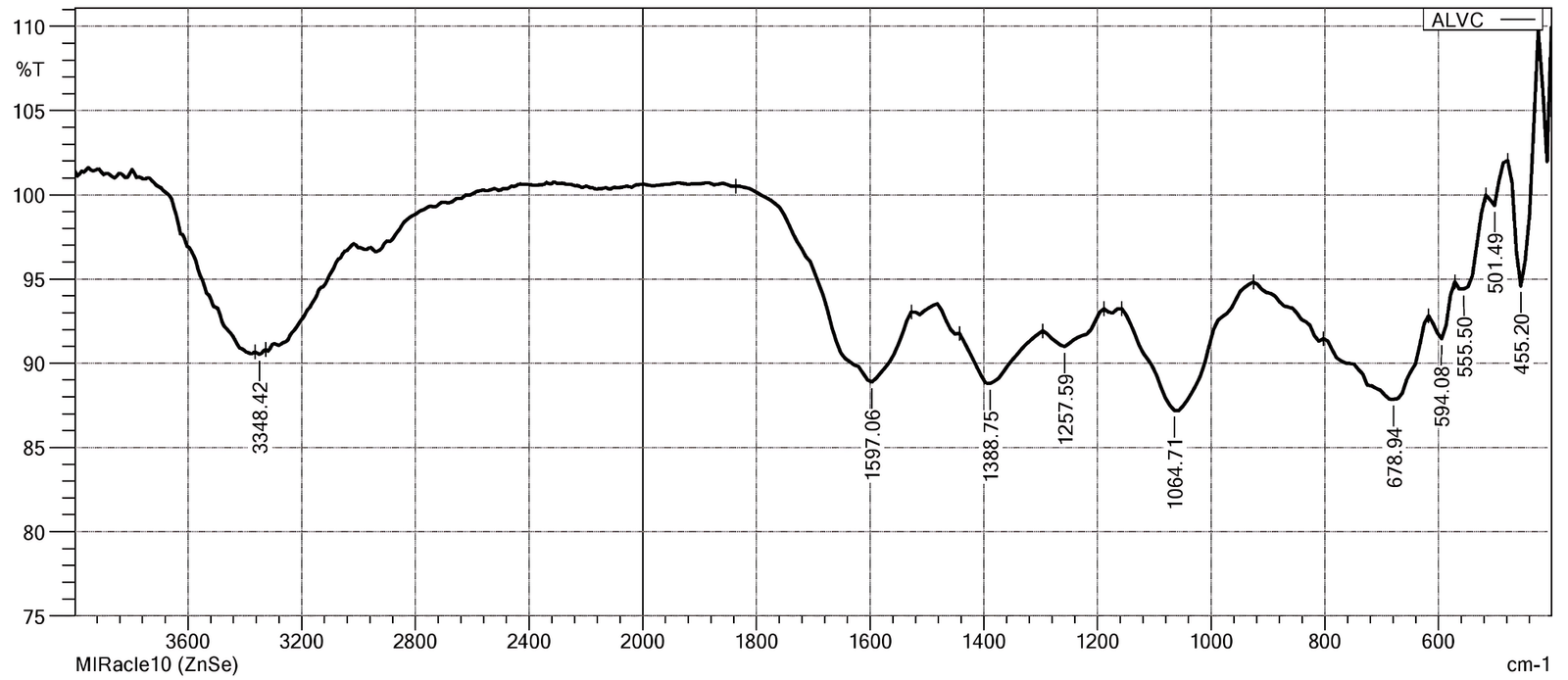
FTIR spectroscopy functions as a robust analytical practice in phytochemical studies by aiding the rapid description of functional group architectures within complex plant matrices, thereby presenting valuable preliminary evidence of secondary metabolite classes and supporting structure-activity interpretations prior to thorough compound-level analyses (42, 43). The FTIR spectrum of V. capparidifolium aqueous extract displayed absorption bands corresponding to various functional groups. A noticeable broad band at ~3348 cm-1 was suggestive of hydrogen-bonded O-H and N-H stretching vibrations correlated with carboxylic acids, phenolic compounds, and amines. Peaks discerned around 1597 cm-1 and 1388 cm-1 were assigned to aromatic C-C stretching, conjugated diene vibrations, and sulfate-related S=O stretching modes. Additional absorptions in the fingerprint region, particularly at ~1258, 1065, and 679 cm-1, parallelled to C-O, C-N, C-F, C-Br, and out-of-plane bending vibrations, establishing the presence of aliphatic, aromatic, and heteroatom-containing functional moieties in the extract (Figure 1, Table 1). FTIR analysis of V. capparidifolium ethanolic leaf extract indicated the presence of polyphenols, carboxylic acids, esters, alkanes, and related bioactive functional groups (22). These functional group signatures are consistent with previous FTIR reports on Tylophora indica (Syn. Vincetoxicum indicum), suggesting the occurrence of flavonoids, terpenoids, and other secondary metabolites associated with pharmacological activity (44, 45).
Metabolic Richness of V. capparidifolium Extract
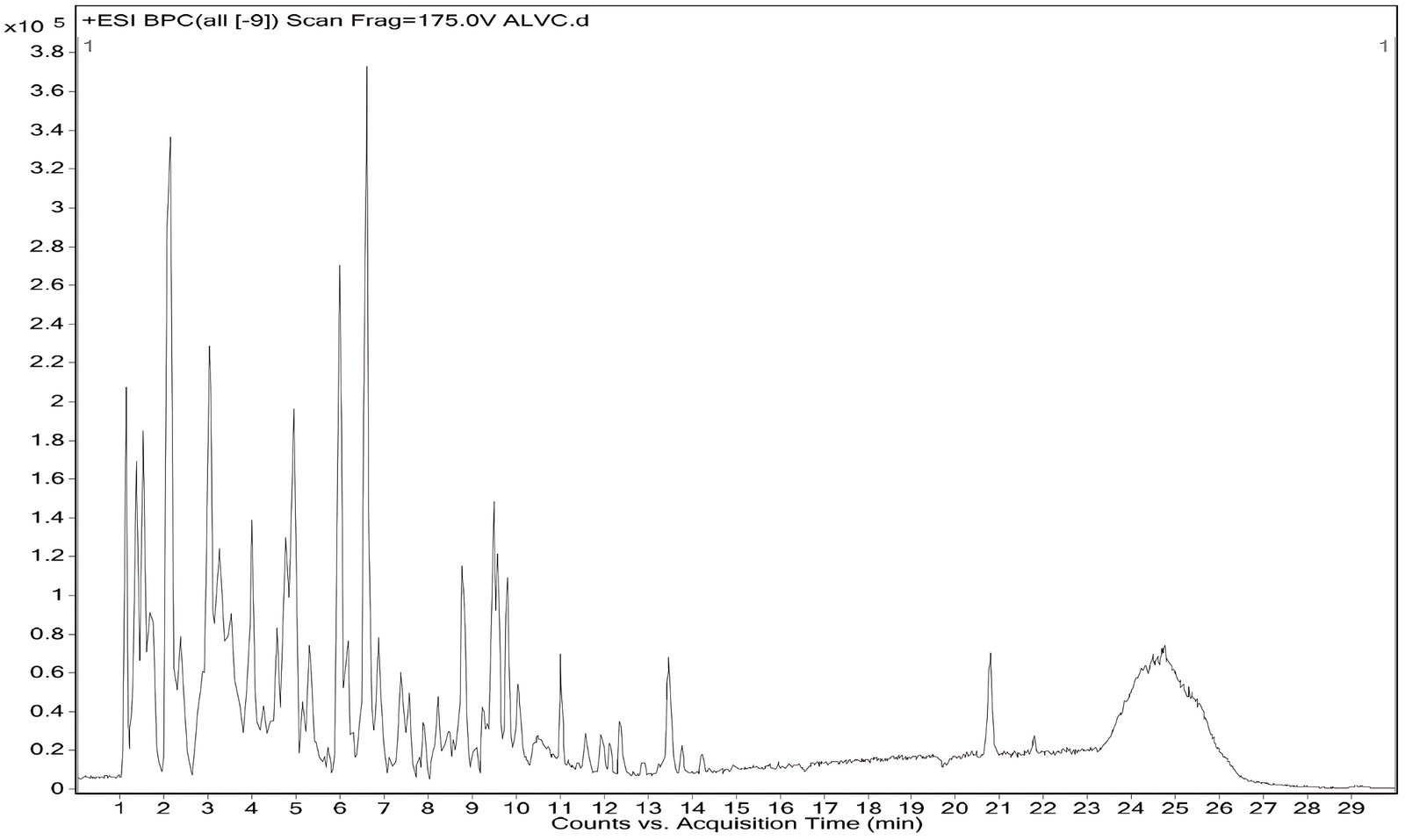
LC-MS analysis is a powerful technique in phytochemical research that enables sensitive separation, identification, and quantification of diverse metabolites in complex plant extracts, thereby facilitating comprehensive chemical profiling and bioactive compound discovery (46). LC-MS profiling of the V. capparidifolium leaf aqueous extract in positive ionization mode enabled the identification of 16 compounds belonging to alkaloids, flavonoid glycosides, flavonols, phenols, peptides, nucleosides, and fatty acid derivatives (Figure 2, Table 2). Major alkaloids, including tylophorine, vincetene, reserpine, and rescinnamine, were detected between RT 1.10-7.42 min, while flavonoids such as quercetin, trifolin, luteolin, and fisetin eluted between RT 3.20-8.56 min; nucleosides and fatty acid derivatives were also observed, indicating marked chemical heterogeneity. All compounds showed match values above 90%, supporting putative metabolite annotation.
| RT | Compound | Compound class | Molecular formula | MW (g/mol) | Match (%) |
|---|---|---|---|---|---|
| 1.10 | Tylophorine | Alkaloid | C24H27NO4 | 393.5 | 98.4 |
| 1.59 | Juglalin | Flavonoid glycoside | C20H18O10 | 418.3 | 96.6 |
| 2.13 | Vincetene | Alkaloid | C22H27NO3 | 353.4 | 99.7 |
| 3.20 | Trifolin | Flavonoid glycoside | C21H20O11 | 448.4 | 97.2 |
| 4.07 | 4-Trifluoromethylphenol | Phenol | C7H5F3O | 162.11 | 90.2 |
| 4.98 | Quercetin | Flavonol | C15H10O7 | 302.23 | 90.8 |
| 5.15 | Leucylproline | Dipeptide | C11H20N2O3 | 228.29 | 92.6 |
| 6.29 | Albine | Tetrahydropyridine | C14H20N2O | 232.32 | 95.1 |
| 6.81 | Rescinnamine | Indole alkaloid | C35H42N2O9 | 634.7 | 90.1 |
| 7.42 | Reserpine | Alkaloid | C33H40N2O9 | 608.7 | 91.3 |
| 8.31 | Hypoxanthine | Oxopurine | C5H4N4O | 136.11 | 96.2 |
| 9.64 | Geranyl 2-ethylbutyrate | Carboxylic ester | C16H28O2 | 252.39 | 90.9 |
| 10.07 | 1-Stearoylglycerol | Long-chain fatty acid alcohol | C21H42O4 | 358.6 | 97.9 |
| 13.64 | Uridine | Ribonucleoside | C9H12N2O6 | 244.20 | 90.1 |
| 20.52 | 3-Methyl-alpha-ionyl acetate | Acetate ester | C16H26O2 | 250.38 | 92.7 |
| 25.14 | Cytidine | Pyrimidine nucleoside | C9H13N3O5 | 243.22 | 91.6 |
| Abbreviations: RT retention time (minutes), MW molecular weight | |||||
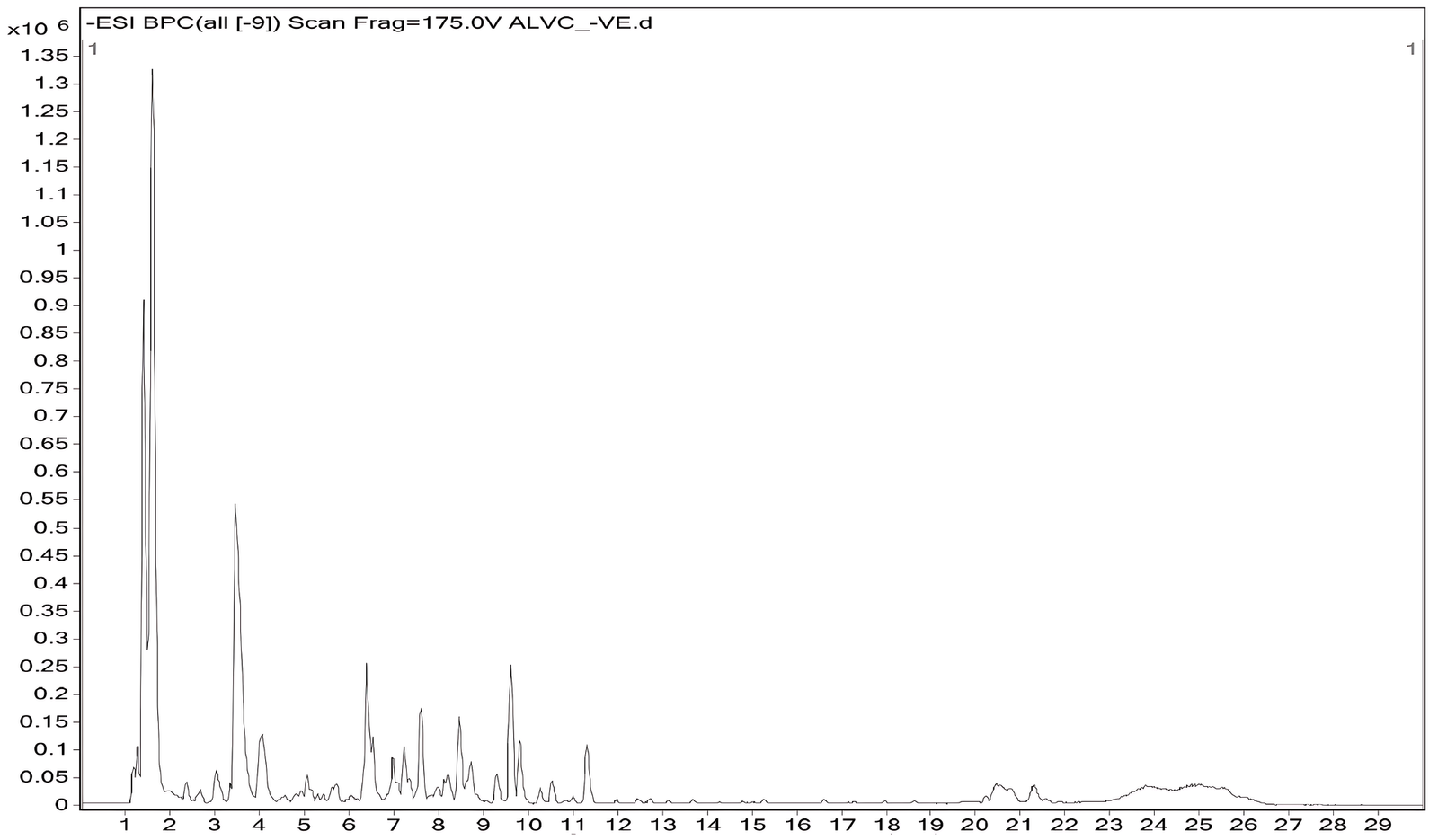
In the negative ionization mode, 12 compounds were detected, highlighting phenolic, flavonoid, and fatty acid amide constituents. Notable compounds included tylophorine, vincetene, catechin, trifolin, apigetrin, quercetin, fisetin, and quercitrin, with retention times spanning 1.33-11.24 min. Fatty acid amides such as oleamide and hexadecanamide and nucleosides like uridine were also identified, indicating the complementarity of negative mode detection for acidic and polar metabolites. Several compounds, particularly tylophorine, vincetene, quercetin, trifolin, and uridine, were detected in both positive and negative modes with minimal variation in retention times. The presence of tylophorine and vincetene in V. capparidifolium is notable, as these alkaloids are recognized chemotaxonomic markers of the genus Vincetoxicum (21, 22, 47). The recurrence of metabolites across both positive and negative ionization modes in the aqueous extract indicates robust ionization behavior of the sample (48). Previous LC-MS studies on T. indica identified several compounds as tylophorinidine derivatives based on database comparisons, while detailed ESI-MSn and LC/MSn analyses of Tylophora atrofolliculata enabled the identification of multiple phenanthroindolizidine alkaloids (49, 50). Comparative LC-MS analysis in positive and negative ionization modes revealed solvent-dependent phytochemical differences, with the aqueous extract mainly enriched in alkaloids, flavonoids, nucleosides, and other metabolites, whereas V. capparidifolium ethanolic extract displayed a wider range of flavonoids, phenolic acids, fatty acids, amides, and structurally diverse secondary metabolites (22). Collectively, dual-mode LC-MS profiling demonstrates the flavonoid- and alkaloid-rich nature of V. capparidifolium leaf extract and highlights its complex, multi-class chemical composition (Figure 3, Table 3) (51).
| RT | Compound | Compound class | Molecular formula | MW (g/mol) | Match (%) |
|---|---|---|---|---|---|
| 1.331 | Tylophorine | Alkaloid | C24H27NO4 | 393.5 | 99.4 |
| 1.501 | Vincetene | Alkaloid | C22H27NO3 | 353.4 | 99.0 |
| 2.862 | Catechin | Phenol | C15H14O6 | 290.27 | 95.5 |
| 3.047 | Rutin | Tetrahydroxy flavone | C27H30O16 | 610.5 | 92.1 |
| 3.624 | Trifolin | Flavonoid glycoside | C21H20O11 | 448.4 | 91.8 |
| 4.119 | Apigetrin | Glycosyloxyflavone | C21H20O10 | 432.4 | 97.6 |
| 5.082 | Luteolin | Flavonoid | C15H10O6 | 286.24 | 94.3 |
| 6.607 | Quercetin | Flavonol | C15H10O7 | 302.23 | 97.2 |
| 7.330 | Uridine | Ribonucleoside | C9H12N2O6 | 244.20 | 90.4 |
| 7.718 | Hexadecanamide | Fatty acid amide | C16H33NO | 255.44 | 90.6 |
| 8.566 | Fisetin | Tetrahydroxy flavone | C15H10O6 | 286.23 | 92.8 |
| 9.82 | Quercitrin | Tetrahydroxy flavone | C21H20O11 | 448.4 | 94.6 |
| 11.244 | Oleamide | Fatty acid amide | C18H35NO | 281.5 | 94.4 |
| Abbreviations: RT retention time (minutes), MW molecular weight. | |||||
Functional Enrichment of Intersecting Targets and Molecular Docking
Network pharmacology provides a systematic framework to detect multi-target interactions and basic mechanisms of bioactive compounds, while gene ontology and enrichment analyses are central for the analytical categorization of genes (52). A total of 94 overlapping genes between tylophorine-associated targets and disease-related genes were subjected to functional enrichment analysis to characterize their biological roles. The selection of genes associated with liver inflammation, diabetic liver disease, and end-stage liver disease was based on the traditional use of V. capparidifolium in the management of jaundice (20, 21). As jaundice commonly arises from hepatic inflammation, metabolic dysfunction, and progressive liver damage, the inclusion of these disease categories provides a broader representation of underlying liver pathophysiology (53). Integrating these disease-associated targets with tylophorine-predicted targets and analyzing their intersection enabled the identification of putative candidate genes potentially associated with the observed biological activities (54).
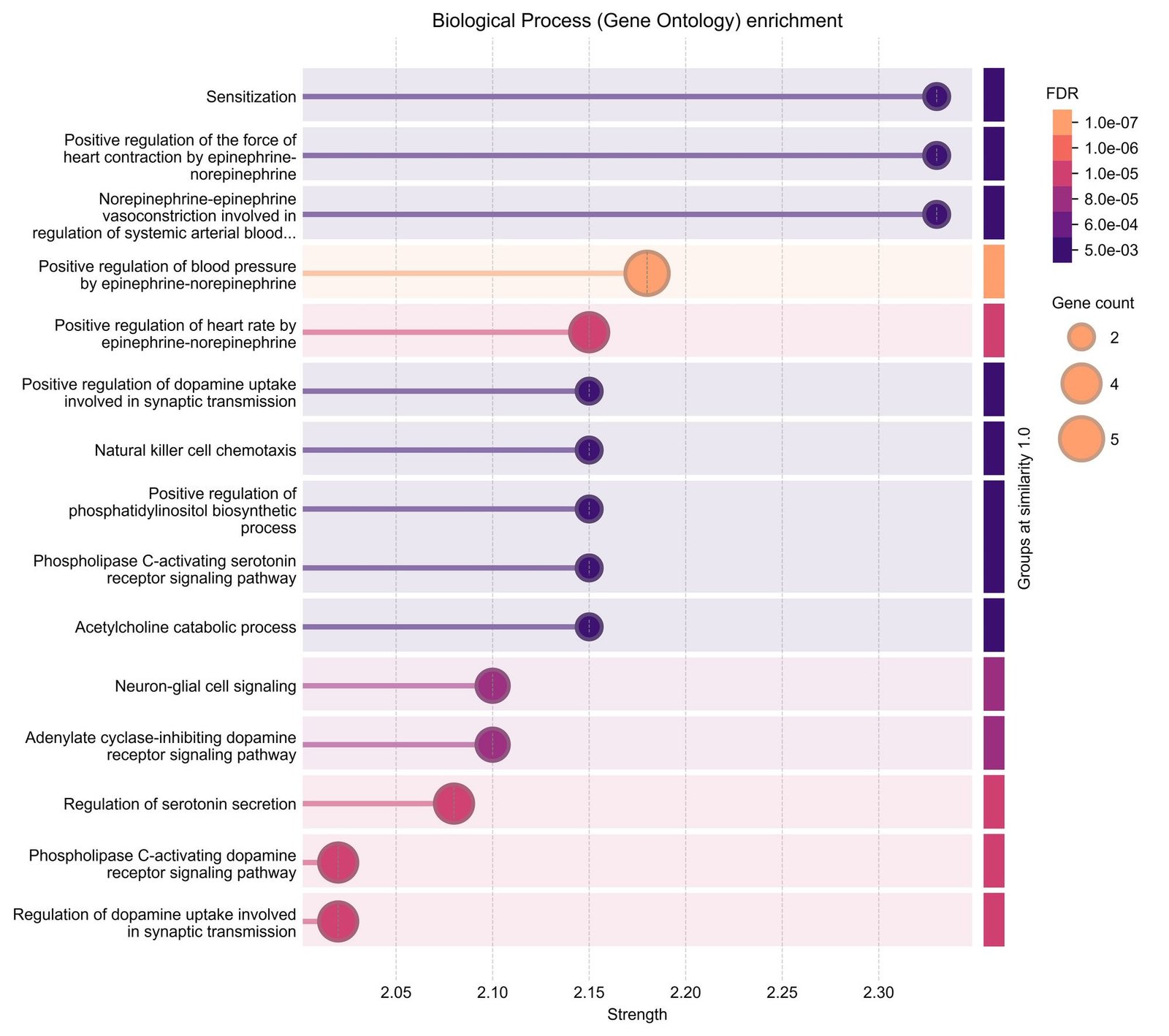
Gene ontology analysis revealed enrichment across three different categories. In the biological processes category, the genes were primarily associated with intracellular signal transduction, regulation of cellular processes, and phosphorylation-related processes. The targets were predominantly localized to intracellular regions such as the cytosol, nucleus, and membrane-associated complexes in the cellular components category. Enriched terms mainly included protein binding, kinase activity, and enzyme binding in terms of molecular functions category (Figure 4). Pathway enrichment analysis further showed that the intersecting genes were enriched in multiple signaling pathways related to cellular regulation and signal transduction.
The protein-protein interaction network constructed from the intersecting genes was analyzed using topological parameters, including degree, closeness, and radiality. Among the top-ranked hub genes identified, MTOR consistently exhibited the highest centrality values across all three parameters, followed by PIK3CA and CHUK, indicating their prominent positions within the network (Figure 5). Other genes, including PIK3CD, PRKCA, PRKCB, and PRKCG, also demonstrated notable connectivity and network influence, while HSP90AB1, CHUK, ESR1, and SLC6A3 showed moderate representation across the topological analyses. A composite scoring system integrating normalized topological parameters and disease relevance was employed to prioritize key hub genes, enabling a balanced evaluation of network significance and biological importance (54, 32). Such multi-criteria tactics are broadly used in network pharmacology to upgrade target selection consistency by combining structural and functional attributes (32, 33). As a result of this approach integrating normalized centrality values and disease relevance, MTOR, PIK3CA, and CHUK were identified as the top-ranked genes (Table 4). Accordingly, the corresponding protein structures, MTOR (PDB ID: 4JSV), PIK3CA (PDB ID: 3HHM), and CHUK (PDB ID: 4KIK), were selected for docking. MTOR, a key regulator of cell growth, metabolism, and autophagy, has been commonly associated in liver pathophysiology, where its dysregulation causes hepatic inflammation, insulin resistance, and progression to fibrosis and hepatocellular carcinoma (55, 56). PIK3CA, encoding the catalytic subunit of phosphatidylinositol 3-kinase, plays a key role in the PI3K-AKT signaling pathway, which regulates glucose metabolism, lipid homeostasis, and cell survival, and is strongly associated with the development of diabetic liver disease and metabolic dysfunction (57, 58). CHUK (IKKα), a critical component of the NF-κB signaling pathway, is involved in regulating inflammatory responses and has been linked to chronic liver inflammation and progression to liver injury through modulation of cytokine-mediated signaling (59, 60).
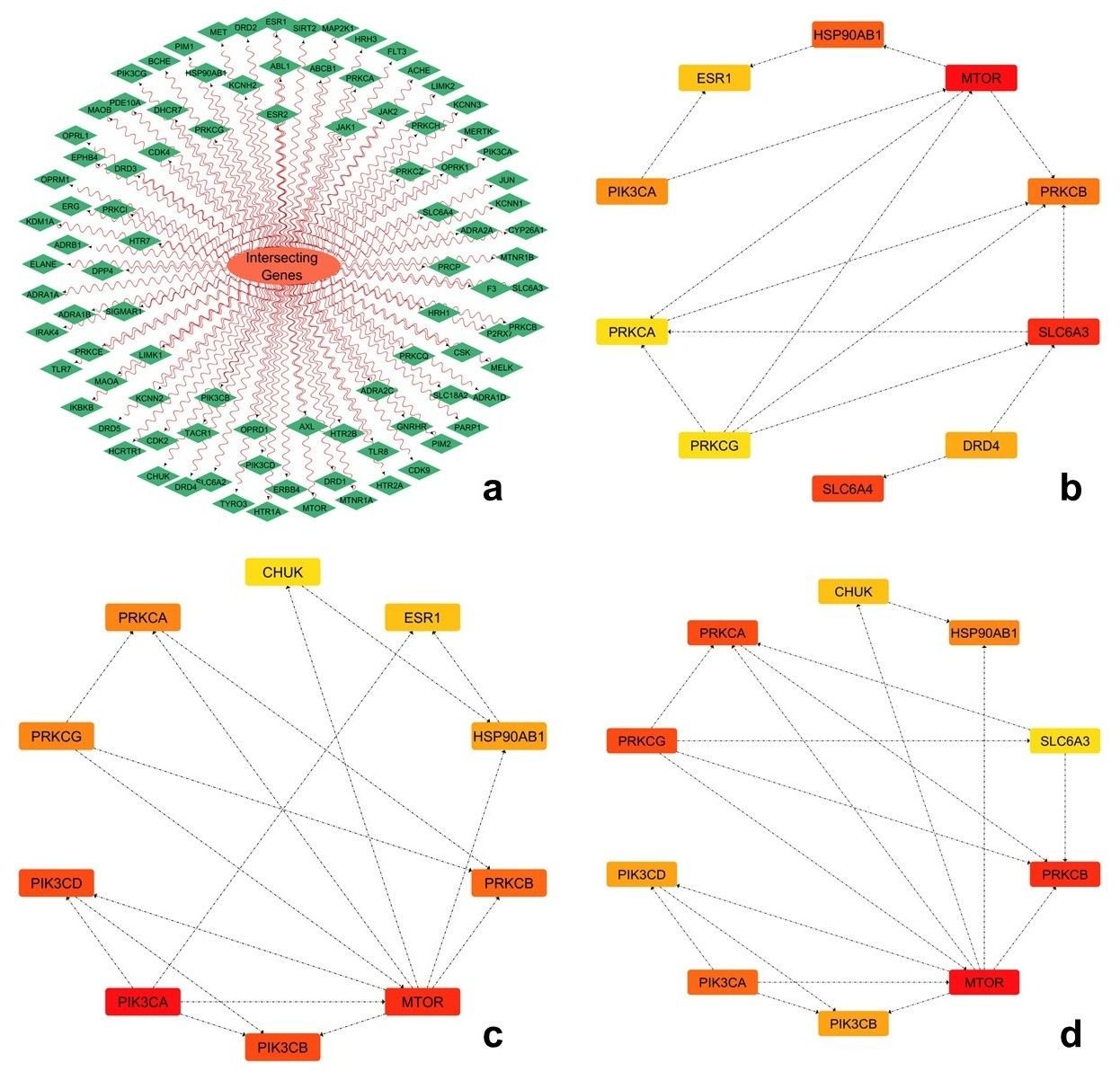
| Gene | Degree | Closeness | Radiality | Disease Score | Final Score |
|---|---|---|---|---|---|
| MTOR | 0.95 | 0.92 | 0.90 | 0.98 | 0.94 |
| PIK3CA | 0.90 | 0.88 | 0.85 | 0.95 | 0.90 |
| CHUK | 0.85 | 0.82 | 0.80 | 0.88 | 0.84 |
| PIK3CB | 0.83 | 0.80 | 0.78 | 0.82 | 0.81 |
| PIK3CD | 0.81 | 0.78 | 0.76 | 0.80 | 0.79 |
| PRKCA | 0.80 | 0.77 | 0.75 | 0.70 | 0.76 |
| PRKCB | 0.78 | 0.75 | 0.73 | 0.68 | 0.74 |
| PRKCG | 0.76 | 0.73 | 0.71 | 0.69 | 0.73 |
| HSP90AB1 | 0.75 | 0.72 | 0.70 | 0.72 | 0.73 |
| ESR1 | 0.72 | 0.70 | 0.68 | 0.69 | 0.70 |
| SLC6A3 | 0.70 | 0.68 | 0.66 | 0.65 | 0.67 |
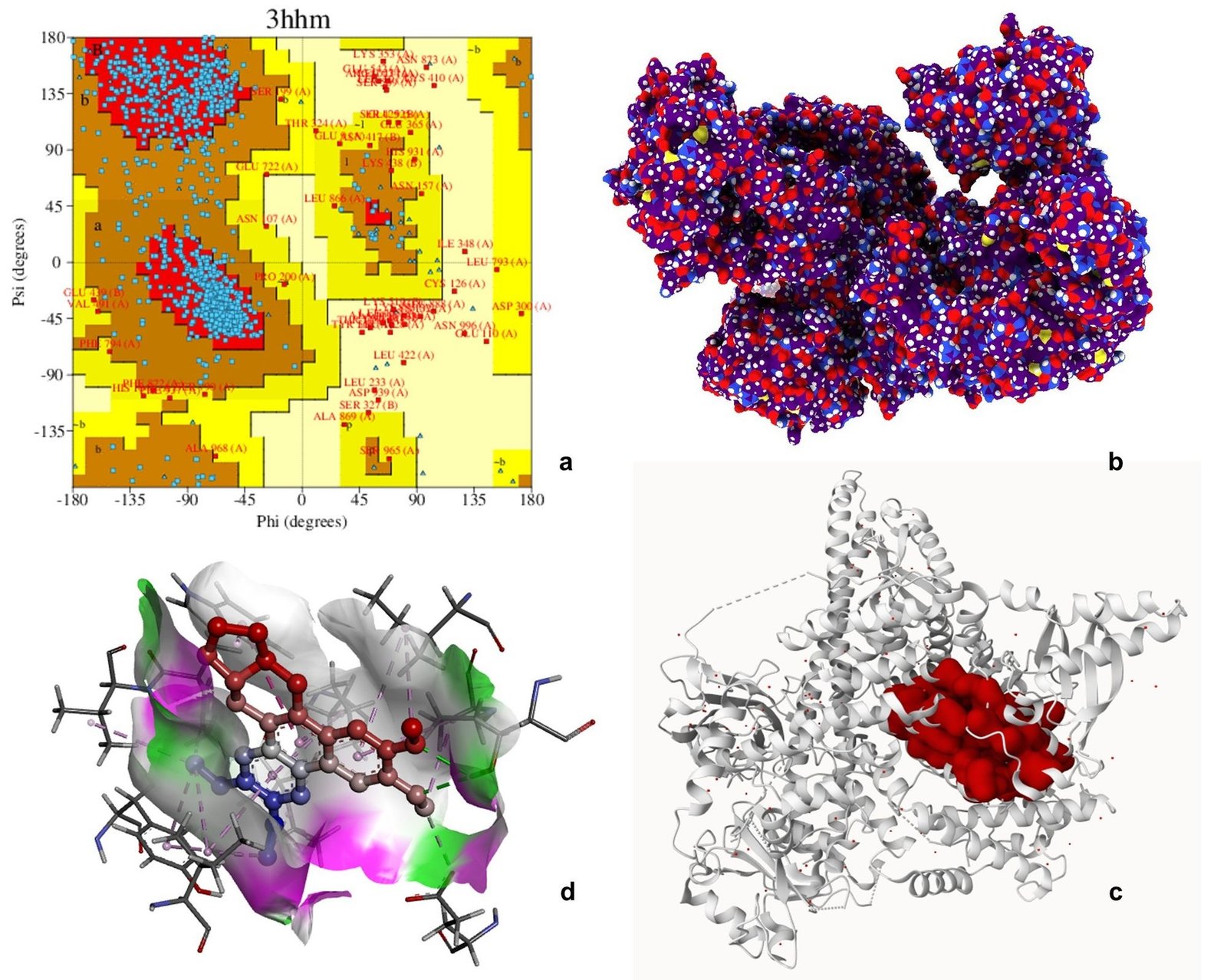
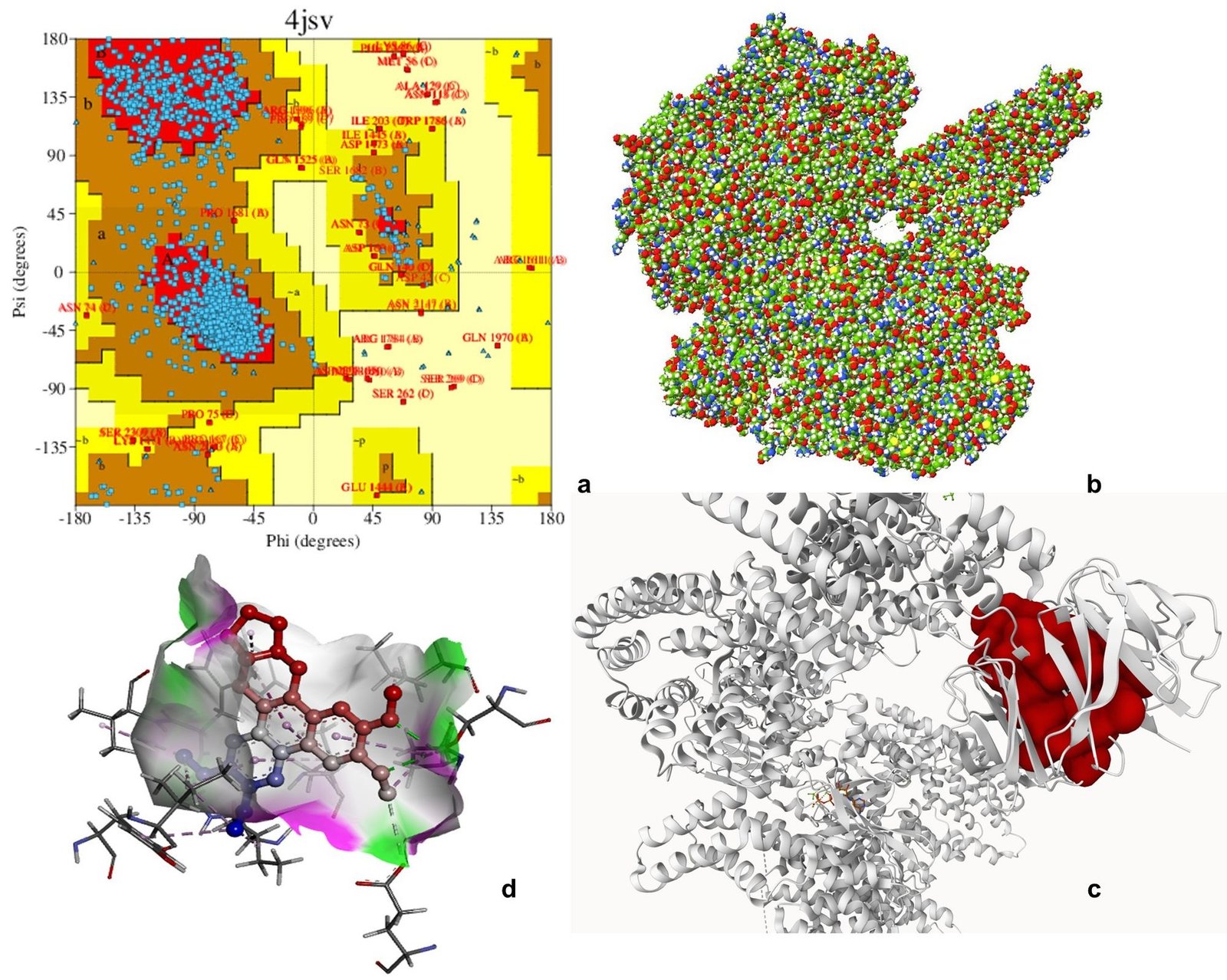
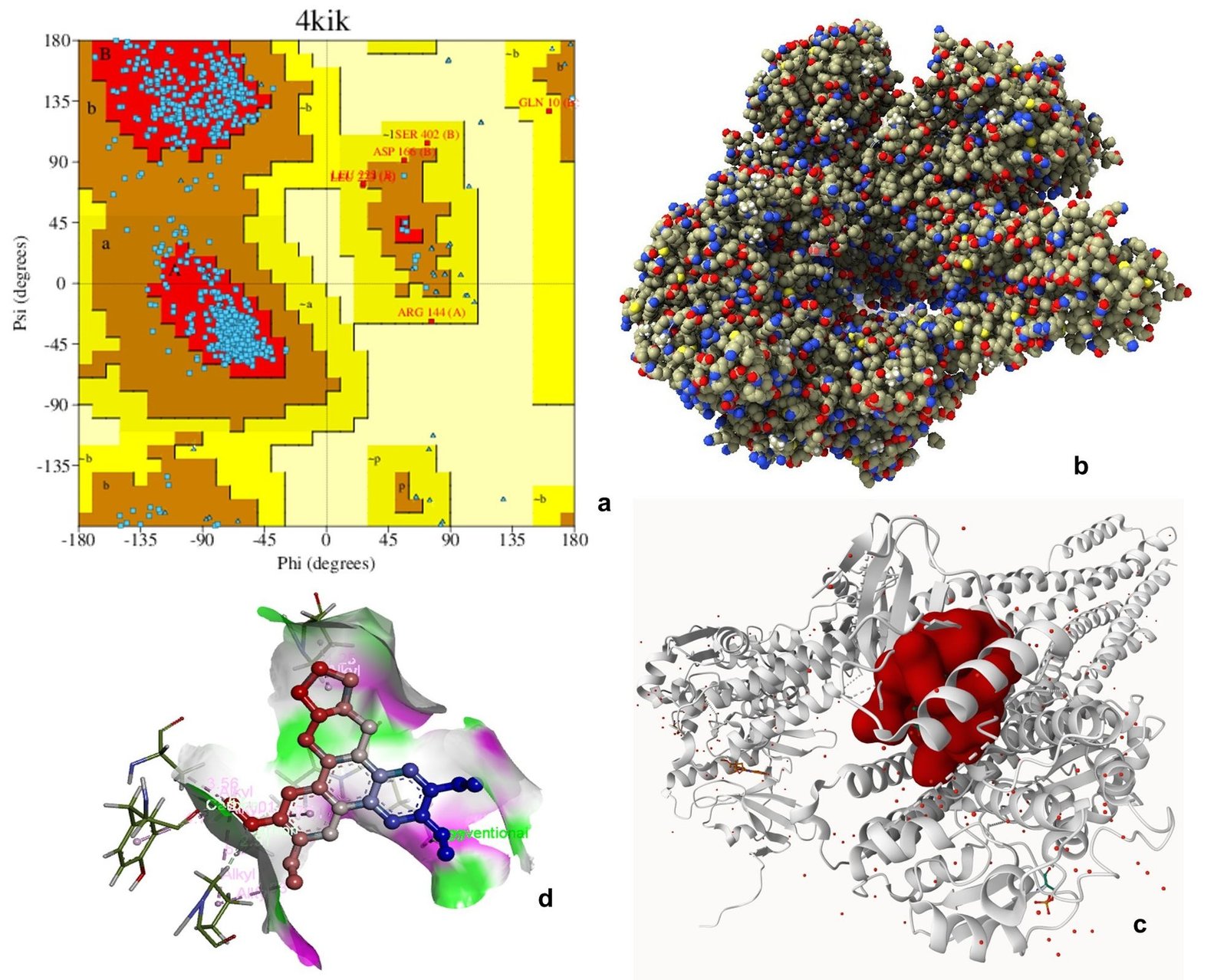
Ramachandran plot analysis of 3HHM, 4JSV, and 4KIK revealed that most residues were clustered in the favored regions (α-helix and β-sheet), with very few residues in disallowed regions, indicating acceptable backbone conformations and good structural quality suitable for downstream computational analysis. The Ramachandran plot is an essential tool for evaluating the stereochemical quality of protein structures by analyzing the distribution of backbone dihedral angles (φ and ψ), thereby helping to identify energetically favorable conformations and structural reliability (61). Molecular docking is widely employed to predict the binding affinity and interaction patterns of bioactive compounds with target proteins, providing insights into their potential mechanism of action at the molecular level (62).
Molecular docking analysis was performed to evaluate the binding affinity of tylophorine with the selected target proteins. The compound exhibited binding interactions with all three proteins, with binding energies of -8.9 kcal mol-1 for 3HHM, -8.1 kcal mol-1 for 4JSV, and -7.9 kcal mol-1 for 4KIK. The interactions involved key active site residues and were predominantly mediated by hydrogen bonding, van der Waals forces, alkyl, and π-related interactions. Among the three targets, 3HHM demonstrated the highest binding affinity. Overall, the docking results suggest favorable binding interactions between tylophorine and the selected target proteins, warranting further experimental validation (Figures 6-8, Table 5). Previous molecular docking studies have shown that tylophorine binds within the ATP-binding pocket of VEGFR2 with an affinity of -7 kcal mol-1, involving key residues such as Lys868, Leu870, His879, Leu882, and Leu912, with hydrophobic interactions predominating over hydrogen bonding (63). Similarly, phenanthrene-based tylophorine derivatives demonstrated favorable binding interactions with the FabH enzyme, supporting their biological activity, with selected compounds exhibiting significant antimicrobial potential and compliance with drug-likeness criteria (64).
| Protein name | PDB ID | Residues Involved | Type of Interactions | H-bonds |
|---|---|---|---|---|
| Phosphatidylinositol 3-kinase (PI3Kα, PIK3CA) | 3HHM | GLN859, LYS802, SER774, ILE800, MET772, TRP780, PRO778, TYR836, ILE848, ILE932 | H-bond, van der Waals, Alkyl, π-alkyl | 3 |
| Serine/threonine-protein kinase MTOR | 4JSV | SER2165, ILE2356, TRP2239, PRO2169, LEU2185, MET2345, TYR2225 | H-bond, π–π stacking, Alkyl, π-alkyl, van der Waals | 1 |
| Inhibitor of nuclear factor kappa-B kinase subunit beta (IKKβ) | 4KIK | GLN1937, PRO1940, LEU1936, ASP2145, TYR2144, ALA1971 | H-bond, Alkyl, π-alkyl, van der Waals | 1 |
Anti-inflammatory Effect of V. capparidifolium Extract
The anti-inflammatory potential of V. capparidifolium leaf aqueous extract was evaluated using the albumin denaturation assay and compared with aspirin as the standard. The extract exhibited protein denaturation over the range of 10-250 µg mL-1, increasing from 9.59% to 68.05%, whereas aspirin showed higher inhibition (20.15-80.36%) (Figure 9). The IC50 value of the extract (153.14 µg mL-1) was higher than that of aspirin (93.91 µg mL-1). The anti-proteinase activity of the extract was assessed using the trypsin inhibition assay. An inhibition of proteolytic activity was observed, with percentage inhibition increasing from 10.33% to 71.76% across the tested concentrations, while aspirin exhibited higher inhibition (20.33-78.90%). The IC50 value of the extract (139.21 µg mL-1) was greater than that of aspirin (96.91 µg mL-1). The membrane stabilizing activity of the extract was evaluated using the heat-induced hemolysis assay. The extract showed protection against erythrocyte lysis, with inhibition increasing from 10.64% to 70.25%, whereas aspirin demonstrated higher values (20.93-80.54%). The IC50 value of the extract (143.70 µg mL-1) was higher than that of aspirin (91.80 µg mL-1). Although the extract exhibited concentration-dependent activity, its potency remained lower than that of aspirin, as reflected by the higher IC50 values.
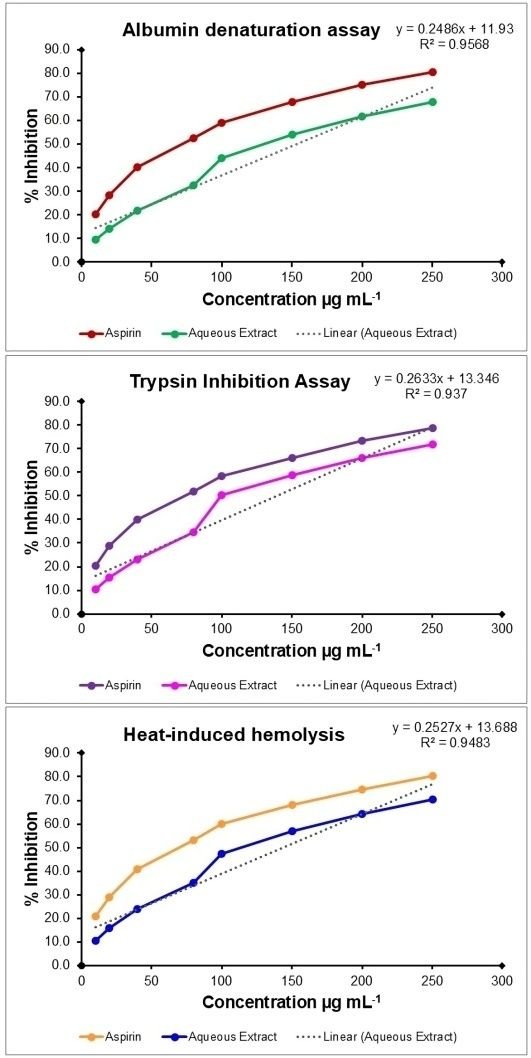
To the best of current knowledge, reports on the in vitro anti-inflammatory activity of Vincetoxicum species are limited. V. arnottianum exhibited significant protection against heat-induced protein denaturation, with ethyl acetate (73%) and methanolic fraction (70%) showing notable inhibition at 500 µg mL-1, comparable to standard drugs diclofenac potassium and aspirin (65). Given that there are few studies on the trypsin inhibitory activity of Vincetoxicum species, the genus demonstrates anti-inflammatory capabilities through cytokine suppression and protein denaturation protection, indicating that similar phytochemicals underpin the reported action (66). V. arnottianum fractions, notably ethyl acetate, exhibited strong in vivo anti-edema effects, linking such in vitro stabilization to broader suppression of oxidative stress and pro-inflammatory mediators such as TNF-α (67). Other members of Apocynaceae, such as Carissa carandas, have also demonstrated anti-inflammatory potential through RBC membrane stabilization, where steroidal saponins from C. carandas leaves exhibited notable in vitro anti-inflammatory properties, preventing hypotonicity-induced hemolysis (68). Additionally, phytoconstituents isolated from C. carandas roots, including lupeol, stigmasterol, oleanolic acid, carissone, and scopoletin, have shown capacity to suppress pro-inflammatory mediators like cytokines and prostaglandins, aligning mechanistically with Vincetoxicum species' membrane-protective effects (69).
Antidiabetic Potential of V. capparidifolium Extract
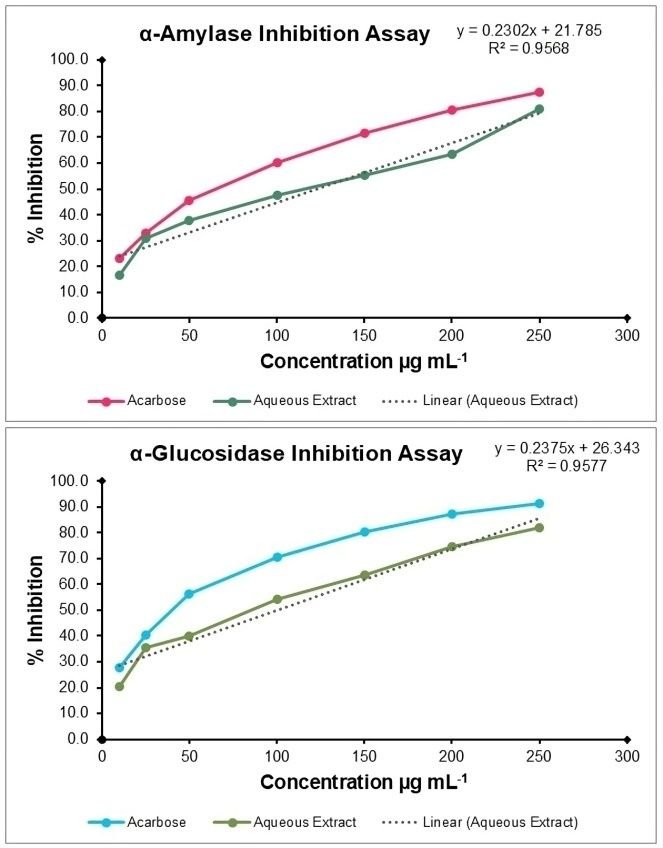
The aqueous extract of V. capparidifolium demonstrated inhibitory effect against both α-amylase and α-glucosidase enzymes over the tested range of 10-250 µg mL-1 (Figure 10). In the α-amylase assay, the extract exhibited inhibition ranging from 16.48% at 10 µg mL-1 to 81.12% at 250 µg mL-1, with an IC50 value of 121.88 µg mL-1. The standard drug acarbose showed comparatively higher inhibitory activity, with inhibition reaching 87.32% at the highest concentration and an IC50 of 83.92 µg mL-1. In the α-glucosidase assay, the extract displayed relatively stronger inhibitory activity, with values increasing from 20.28% at 10 µg mL-1 to 81.75% at 250 µg mL-1. The IC50 value was determined to be 99.61 µg mL-1. Acarbose exhibited greater potency, with an IC50 of 53.29 µg mL-1 and a maximum inhibition of 91.48%. Notably, the extract demonstrated comparatively higher efficacy against α-glucosidase than α-amylase. Despite exhibiting dose-dependent inhibition of both enzymes, the extract was less potent than acarbose, as indicated by its higher IC50 values. Several members of this family exhibit notable antidiabetic potential; for instance, alkaloids from Catharanthus roseus have been reported to reduce blood glucose levels and improve glucose tolerance, while Pterocarpus marsupium bark extract has demonstrated hypoglycemic effects through enhanced insulin secretion and improved insulin sensitivity (19).
Cytotoxic and Apoptotic Effects of V. capparidifolium Extract on HepG2 Cells
The cytotoxic potential of V. capparidifolium aqueous extract was evaluated using the MTT assay (Figure 11a). Treatment with increasing concentrations of the sample resulted in a marked decline in viable cells, with an IC50 value of 168.04 µg mL-1. To further confirm the mode of cell death, AO/EtBr dual staining was performed (Figure 11b-d). Control cells exhibited uniformly green fluorescence with intact morphology, indicating viable cells (Figure 11b). In contrast, doxorubicin-treated cells showed pronounced red/orange fluorescence with nuclear condensation and membrane damage, characteristic of apoptotic cell death (Figure 11c). Cells treated with the sample at 168.04 µg mL-1 concentration also displayed increased red/orange fluorescence along with reduced green fluorescence, suggesting the induction of apoptosis (Figure 11d). Additionally, DNA fragmentation analysis revealed intact genomic DNA in control cells (Figure 11e), whereas cells treated with the extract at IC50 concentration showed fragmented DNA patterns (Figure 11f), further confirming apoptosis-mediated cytotoxicity. Collectively, these findings indicate that the sample exerts antiproliferative activity by inducing apoptotic cell death.
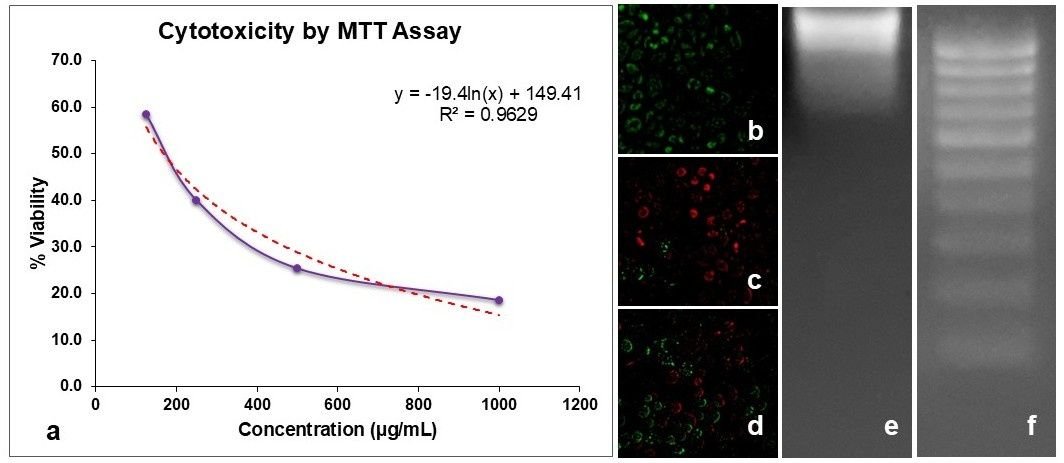
Previous studies on the genus Vincetoxicum have demonstrated notable anticancer potential through MTT-based cytotoxicity assays. The ethanolic leaf extract of V. capparidifolium exhibited dose-dependent cytotoxicity against HepG2 hepatocellular carcinoma cells, reducing cell viability to 18% at 1000 µg mL-1 with an estimated IC50 value of 163 µg mL-1. Apoptotic cell death was supported by AO/EtBr staining, which revealed membrane compromise and nuclear condensation, and by DNA fragmentation analysis showing a characteristic ladder pattern, indicative of apoptosis rather than necrosis (19). The methylene chloride fraction of V. pumilum exhibited potent cytotoxicity against leukemic cell lines HL-60 and K562, with IC50 values of 8.3 and 12.5 µg mL-1, respectively, and induced apoptosis through sub-G1 arrest and upregulation of pro-apoptotic markers, while showing minimal toxicity toward normal lymphocytes (69). Similarly, the methanolic extract of V. arnottianum markedly reduced cell viability (10-30%) and inhibited invasion by up to 99% in pediatric alveolar rhabdomyosarcoma (RH-30) cells at 25 µg mL-1 (65). The MTT assay conducted on V. hirundinaria extracts demonstrated that extracts exerted no cytotoxic effects on HaCaT cells at any tested concentration (0.25-1 mg mL-1), indicating a favorable safety profile toward normal cells (70).
Conclusion
The aqueous leaf extract of V. capparidifolium showed a chemically diverse phytochemical profile, characterized by FTIR-identified functional groups and LC-MS-detected alkaloids, flavonoids, nucleosides, and fatty acid derivatives. Network pharmacology and molecular docking analyses identified potential molecular targets associated with liver-related disorders and suggested favorable interactions of tylophorine with selected proteins. The extract demonstrated concentration-dependent anti-inflammatory, anti-diabetic, cytotoxic, and apoptotic effects in vitro. As the present findings are limited to in silico and in vitro analyses, further studies are required to validate the biological efficacy and safety of the extract.
Declarations
Acknowledgment
The authors express their sincere gratitude to the research scholars and staff members of the PG and Research Department of Botany, Kongunadu Arts and Science College (Autonomous), Coimbatore, for their constant support, cooperation, and encouragement throughout the course of this research work.
Conflict of Interest
The authors declare no conflict of interest.
Data Availability
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
Ethics Statement
Not applicable.
Funding Information
This work was funded by Council of Scientific and Industrial Research-Human Resource Development Group (CSIR-HRDG), Government of India, through the Direct Senior Research Fellowship (SRF) scheme under grant number (08/1367(25829)/2025-EMR-I).
References
- Galicia-Moreno M, Silva-Gomez JA, Lucano-Landeros S, Santos A, Monroy-Ramirez HC, Armendariz-Borunda J. Liver Cancer: Therapeutic Challenges and the Importance of Experimental Models. Canadian Journal of Gastroenterology and Hepatology. 2021;2021(1):1-10. doi: https://doi.org/10.1155/2021/8837811
- Zheng J, Wang S, Xia L, Sun Z, Chan KM, Bernards R, et al. Hepatocellular carcinoma: signaling pathways and therapeutic advances. Sig Transduct Target Ther. 2025;10(1):1-43. doi: https://doi.org/10.1038/s41392-024-02075-w
- Masuzaki R. Liver Cancer: Improving Standard Diagnosis and Therapy. Cancers. 2023;15(18):4602. doi: https://doi.org/10.3390/cancers15184602
- Anand U, Dey A, Chandel AKS, Sanyal R, Mishra A, Pandey DK, et al. Cancer chemotherapy and beyond: Current status, drug candidates, associated risks and progress in targeted therapeutics. Genes & Diseases. 2023;10(4):1367-1401. doi: https://doi.org/10.1016/j.gendis.2022.02.007
- Zafar A, Khatoon S, Khan MJ, Abu J, Naeem A. Advancements and limitations in traditional anti-cancer therapies: a comprehensive review of surgery, chemotherapy, radiation therapy, and hormonal therapy. Discov Onc. 2025;16(1):607. doi: https://doi.org/10.1007/s12672-025-02198-8
- Liu B, Zhou H, Tan L, Siu KTH, Guan XY. Exploring treatment options in cancer: tumor treatment strategies. Sig Transduct Target Ther. 2024;9(1):1-44. doi: https://doi.org/10.1038/s41392-024-01856-7
- Dzobo K. The Role of Natural Products as Sources of Therapeutic Agents for Innovative Drug Discovery. Pradiningsih A, Hasan YM, Nopitasari BL, Adikusuma W. Potential Occurrence of Drug related problems (DRPs) in Type 2 Diabetes Mellitus patients. Res J Pharm Technol. 2025;18(4):1597–603. Amsterdam, Netherlands: Elsevier; 2022. doi: https://doi.org/10.1016/b978-0-12-820472-6.00041-4
- Chaachouay N, Zidane L. Correction: Chaachouay, N.; Zidane, L. Plant-Derived Natural Products: A Source for Drug Discovery and Development. Drugs Drug Candidates 2024, 3, 184–207. Ddc. 2026;5(1):14. doi: https://doi.org/10.3390/ddc5010014
- Chidananda C, Thakur G, Datta D, Popat K. A comprehensive review of alkaloids in cancer therapy: focusing on molecular mechanisms and synergistic potential of piperine in colorectal cancer. 3 Biotech. 2025;15(11):403. doi: https://doi.org/10.1007/s13205-025-04535-8
- Hasnat H, Shompa SA, Islam MM, Alam S, Richi FT, Emon NU, et al. Flavonoids: A treasure house of prospective pharmacological potentials. Heliyon. 2024;10(6):e27533. doi: https://doi.org/10.1016/j.heliyon.2024.e27533
- Sun W, Shahrajabian MH. Therapeutic Potential of Phenolic Compounds in Medicinal Plants—Natural Health Products for Human Health. Molecules. 2023;28(4):1845. doi: https://doi.org/10.3390/molecules28041845
- Janardhana SK, Nekrakalaya B. Ethnobotany to bioprospecting of medicinal plants from Western Ghats, India – A review. Ijtk. 2025;24(1):63-72. doi: https://doi.org/10.56042/ijtk.v24i1.16335
- Prameela A, Krishnasamy T. An Investigation on the Age-Old Therapeutics Practiced by the Muthuvan Tribe of Malappuram District, Kerala, India. Cham: Springer Nature Switzerland; 2026. doi: https://doi.org/10.1007/978-3-031-91945-9_21
- Revathi P, Prameela A, Malavika J, Krishnasamy T. Ethnobotanical Studies on Kattunaickan Tribes of Janakikkadu, Kozhikode District, Kerala, India. Cham: Springer Nature Switzerland; 2026. doi: https://doi.org/10.1007/978-3-031-91945-9_23
- Radhakrishnan A, Sivakumar T, Habibulla AK. Deciphering the bioactive potential of Monosis travancorica: Phytochemical profiling, Drug-likeness insights, and Cyclooxygenase-2 targeting docking studies. Ijbb. 2026;63(4):426-441. doi: https://doi.org/10.56042/ijbb.v63i4.17420
- Patil RH, Patil MP, Maheshwari VL. Apocynaceae Plants. Singapore: Springer Nature Singapore; 2023. doi: https://doi.org/10.1007/978-981-99-5406-3
- Patil RH, Patil MP, Maheshwari VL. Traditional Knowledge, Ethnomedicinal Importance and Practical Uses of Apocynaceae Members. Singapore: Springer Nature Singapore; 2023. doi: https://doi.org/10.1007/978-981-99-5406-3_2
- Patil RH, Patil MP, Maheshwari VL. Phytochemistry of Apocynaceae Members. Singapore: Springer Nature Singapore; 2023. doi: https://doi.org/10.1007/978-981-99-5406-3_7
- Patil RH, Patil MP, Maheshwari VL. Biological and Pharmacological Properties of Apocynaceae Members. Singapore: Springer Nature Singapore; 2023. doi: https://doi.org/10.1007/978-981-99-5406-3_8
- Hebbar SS, Harsha H, Hegde R, Shripathi V. Ethno-Medicobotanical Survey in Dharwad-Plants Used for Jaundice. Nelumbo. 2024;46(1):268-272. doi: https://doi.org/10.20324/nelumbo/v46/2004/74169
- Prameela A, Krishnasamy T, D. SS. Ethnobotany, phylogeny, phytochemistry and pharmacological applications of genus Vincetoxicum (Apocynaceae). Discov. Plants. 2024;1(1):1-18. doi: https://doi.org/10.1007/s44372-024-00031-0
- Prameela A, Krishnasamy T, Thangavelu M. Comprehensive metabolite profiling, antioxidant potential and anticancer mechanism of the Indian medicinal plant species Vincetoxicum capparidifolium , Apocynaceae. Journal of Herbs, Spices & Medicinal Plants. 2026;32(3):297-323. doi: https://doi.org/10.1080/10496475.2026.2625097
- Thangaraj P. Quantification of Secondary Metabolites. Cham: Springer International Publishing; 2015. doi: https://doi.org/10.1007/978-3-319-26811-8_8
- Sahraoui A, Hamlaoui M, Chikhi S, Harrat S, Baghriche O, Zertal A, et al. Green synthesis and characterization of titanium dioxide nanoparticles using Eucalyptus globulus leaf extract: Impacts of the mild thermal treatment. Materials Today Sustainability. 2025;31:101193. doi: https://doi.org/10.1016/j.mtsust.2025.101193
- Rafi M, Hayati F, Umar AH, Septaningsih DA, Rachmatiah T. LC-HRMS-based metabolomics to evaluate the phytochemical profile and antioxidant capacity of Cosmos caudatus with different extraction methods and solvents. Arabian Journal of Chemistry. 2023;16(9):105065. doi: https://doi.org/10.1016/j.arabjc.2023.105065
- Stelzer G, Rosen N, Plaschkes I, Zimmerman S, Twik M, Fishilevich S, et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. CP in Bioinformatics. 2016;54(1):1.30.1-1.30.33. doi: https://doi.org/10.1002/cpbi.5
- Gfeller D, Grosdidier A, Wirth M, Daina A, Michielin O, Zoete V. SwissTargetPrediction: a web server for target prediction of bioactive small molecules. Nucleic Acids Research. 2014;42(W1):W32-W38. doi: https://doi.org/10.1093/nar/gku293
- Mering CV. STRING: a database of predicted functional associations between proteins. Nucleic Acids Research. 2003;31(1):258-261. doi: https://doi.org/10.1093/nar/gkg034
- Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, et al. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003;13(11):2498-2504. doi: https://doi.org/10.1101/gr.1239303
- Mukherjee PK, Banerjee S, Kar A. Molecular combination networks in medicinal plants: understanding synergy by network pharmacology in Indian traditional medicine. Phytochem Rev. 2021;20(4):693-703. doi: https://doi.org/10.1007/s11101-020-09730-4
- Gonçalves JP, Francisco AP, Moreau Y, Madeira SC. Interactogeneous: Disease Gene Prioritization Using Heterogeneous Networks and Full Topology Scores. PLoS ONE. 2012;7(11):e49634. doi: https://doi.org/10.1371/journal.pone.0049634
- Razaghi-Moghadam Z, Abdollahi R, Goliaei S, Ebrahimi M. HybridRanker: Integrating network topology and biomedical knowledge to prioritize cancer candidate genes. Journal of Biomedical Informatics. 2016;64:139-146. doi: https://doi.org/10.1016/j.jbi.2016.10.003
- Browne F, Wang H, Zheng H. A computational framework for the prioritization of disease-gene candidates. BMC Genomics. 2015;16(S9):1-10. doi: https://doi.org/10.1186/1471-2164-16-s9-s2
- Dallakyan S, Olson AJ. Small-Molecule Library Screening by Docking with PyRx. New York, NY: Springer New York; 2014. doi: https://doi.org/10.1007/978-1-4939-2269-7_19
- Iqbal D, Alsaweed M, Jamal QMS, Asad MR, Rizvi SMD, Rizvi MR, et al. Pharmacophore-Based Screening, Molecular Docking, and Dynamic Simulation of Fungal Metabolites as Inhibitors of Multi-Targets in Neurodegenerative Disorders. Biomolecules. 2023;13(11):1613. doi: https://doi.org/10.3390/biom13111613
- M A, I MA, Ramalingam K, S R. Evaluation of the Anti-inflammatory, Antimicrobial, Antioxidant, and Cytotoxic Effects of Chitosan Thiocolchicoside-Lauric Acid Nanogel. Cureus. 2023;15(9):e46003. doi: https://doi.org/10.7759/cureus.46003
- Mohan M, Kozhithodi S, Nayarisseri A, Elyas KK. Screening, Purification and Characterization of Protease Inhibitor from Capsicum frutescens. Bioinformation. 2018;14(06):285-293. doi: https://doi.org/10.6026/97320630014285
- Moualek I, Iratni Aiche G, Mestar Guechaoui N, Lahcene S, Houali K. Antioxidant and anti-inflammatory activities of Arbutus unedo aqueous extract. Asian Pacific Journal of Tropical Biomedicine. 2016;6(11):937-944. doi: https://doi.org/10.1016/j.apjtb.2016.09.002
- Hullatti K, Telagari M. In-vitro α-amylase and α-glucosidase inhibitory activity of Adiantum caudatum Linn. and Celosia argentea Linn. extracts and fractions. Indian J Pharmacol. 2015;47(4):425. doi: https://doi.org/10.4103/0253-7613.161270
- Ala AA, Olotu BB, Ohia CMD. Assessment of cytotoxicity of leaf extracts of Andrographis paniculata and Aspilia africana on murine cells in vitro. Arch. Basic Appl. Med. 2018;6(1):61-65. PMID: 30234147; PMCID: PMC6141048
- Ifeoluwa Okeleye B, Mkwetshana NT, Ndip RN. In vitro assessment of the antiproliferative and apoptotic potential of the ethyl acetate extract of Peltophorum africanum on different cancer cell lines. Iran J. Pharm. Res. 2017;16(2):714-724. PMID: 28979326; PMCID: PMC5603881
- Aguree S, Abagale SA, Sackey I. GC–MS and FTIR Characterization of Bioactive Compounds in Aqueous Extracts of Terminalia laxiflora Root and Mitragyna inermis Stem Bark. Journal of Chemistry. 2025;2025(1):1-9. doi: https://doi.org/10.1155/joch/7537345
- Pasieczna-Patkowska S, Cichy M, Flieger J. Application of Fourier Transform Infrared (FTIR) Spectroscopy in Characterization of Green Synthesized Nanoparticles. Molecules. 2025;30(3):684. doi: https://doi.org/10.3390/molecules30030684
- Vanitha A, Vijayakumar S, Ranjitha V, Kalimuthu K. Phytochemical screening and antimicrobial activity of wild and tissue cultured plant extracts of Tylophora indica. Asian J Pharm Pharmacol. 2018;5(1):21-32. doi: https://doi.org/10.31024/ajpp.2019.5.1.3
- Srinivasan K. FT-IR and EDS analysis of the seaweeds Sargassum wightii (brown algae) and Gracilaria corticata (red algae). Int. J. Curr. Microbiol. Appl. Sci. 2014;3(4):341-351. https://www.ijcmas.com/vol-3-4/S.Kannan.pdf
- D’Urso G, Capuano A, Fantasma F, Chini MG, De Felice V, Saviano G, et al. The Role of LC-MS in Profiling Bioactive Compounds from Plant Waste for Cosmetic Applications: A General Overview. Plants. 2025;14(15):2284. doi: https://doi.org/10.3390/plants14152284
- Prameela A, Radhakrishnan A, Krishnasamy T. Elucidating Neuropharmacological Implications of Vincetene: A Multi-target Computational Study on Ataxia, Encephalitis, and Meningitis. Prospect. Pharm. Sci. 2025;23(4):103-114. doi: https://doi.org/10.56782/pps.490
- Mukherjee PK, Banerjee S, Kar A. Molecular combination networks in medicinal plants: understanding synergy by network pharmacology in Indian traditional medicine. Phytochem Rev. 2021;20(4):693-703. doi: https://doi.org/10.1007/s11101-020-09730-4
- Manikkoth S, B D, Sequeira M, Elizabeth Joy A, Rodrigues R. Assessment of brain dopamine levels to evaluate the role of Tylophora indicaethanolic extract on alcohol induced anxiety in Wistar albino rats. Jyp. 2016;8(2):91-95. doi: https://doi.org/10.5530/jyp.2016.2.6
- Cui L, Abliz Z, Xia M, Zhao L, Gao S, He W, et al. On‐line identification of phenanthroindolizidine alkaloids in a crude extract from Tylophora atrofolliculata by liquid chromatography combined with tandem mass spectrometry. Rapid Comm Mass Spectrometry. 2003;18(2):184-190. doi: https://doi.org/10.1002/rcm.1297
- de Jonge NF, Chekmeneva E, Schmid R, Joas D, Truong LJ, van der Hooft JJJ, et al. Cross ionization mode chemical similarity prediction between tandem mass spectra in metabolomics. Nat Commun. 2026;17(1):1-12. doi: https://doi.org/10.1038/s41467-026-69083-y
- Bhattacharjee B, M. Jayadeepa R, Talambedu U, Banerjee S, Joshi J, Princy Mole J, et al. Complex Network and Gene Ontology in Pharmacology Approaches:Mapping Natural Compounds on Potential Drug Target Colon Cancer Network. Cbio. 2011;6(1):44-52. doi: https://doi.org/10.2174/157489311795222400
- Chen HL, Wu SH, Hsu SH, Liou BY, Chen HL, Chang MH. Jaundice revisited: recent advances in the diagnosis and treatment of inherited cholestatic liver diseases. J Biomed Sci. 2018;25(1):75. doi: https://doi.org/10.1186/s12929-018-0475-8
- Gonçalves JP, Francisco AP, Moreau Y, Madeira SC. Interactogeneous: Disease Gene Prioritization Using Heterogeneous Networks and Full Topology Scores. PLoS ONE. 2012;7(11):e49634. doi: https://doi.org/10.1371/journal.pone.0049634
- Wang H, Liu Y, Wang D, Xu Y, Dong R, Yang Y, et al. The Upstream Pathway of mTOR-Mediated Autophagy in Liver Diseases. Cells. 2019;8(12):1597. doi: https://doi.org/10.3390/cells8121597
- Ferrín G, Guerrero M, Amado V, Rodríguez-Perálvarez M, De la Mata M. Activation of mTOR Signaling Pathway in Hepatocellular Carcinoma. Ijms. 2020;21(4):1266. doi: https://doi.org/10.3390/ijms21041266
- Ma C, Yang X, Zhang L, Zhang J, Zhang Y, Hu X. BRCA1 regulates glucose and lipid metabolism in diabetes mellitus with metabolic dysfunction-associated steatotic liver disease via the PI3K/Akt signaling pathway. PLoS ONE. 2025;20(3):e0318696. doi: https://doi.org/10.1371/journal.pone.0318696
- Tian LY, Smit DJ, Jücker M. The Role of PI3K/AKT/mTOR Signaling in Hepatocellular Carcinoma Metabolism. Ijms. 2023;24(3):2652. doi: https://doi.org/10.3390/ijms24032652
- Xiong Y, Torsoni AS, Wu F, Shen H, Liu Y, Zhong X, et al. Hepatic NF-kB-inducing kinase (NIK) suppresses mouse liver regeneration in acute and chronic liver diseases. eLife. 2018;7:e34152. doi: https://doi.org/10.7554/elife.34152
- Sun B, Karin M. NF-κB signaling, liver disease and hepatoprotective agents. Oncogene. 2008;27(48):6228-6244. doi: https://doi.org/10.1038/onc.2008.300
- Park SW, Lee BH, Song SH, Kim MK. Revisiting the Ramachandran plot based on statistical analysis of static and dynamic characteristics of protein structures. Journal of Structural Biology. 2023;215(1):107939. doi: https://doi.org/10.1016/j.jsb.2023.107939
- Agu PC, Afiukwa CA, Orji OU, Ezeh EM, Ofoke IH, Ogbu CO, et al. Molecular docking as a tool for the discovery of molecular targets of nutraceuticals in diseases management. Sci Rep. 2023;13(1):13398. doi: https://doi.org/10.1038/s41598-023-40160-2
- Saraswati S, Kanaujia PK, Kumar S, Kumar R, Alhaider AA. Tylophorine, a phenanthraindolizidine alkaloid isolated from Tylophora indica exerts antiangiogenic and antitumor activity by targeting vascular endothelial growth factor receptor 2–mediated angiogenesis. Mol Cancer. 2013;12(1):82. doi: https://doi.org/10.1186/1476-4598-12-82
- Thongolla R, Pulabala R, Gogula SS, Shenoy TN, Salam AAA, Vankadari SR, et al. Design, synthesis, and molecular docking studies of new phenanthrene-linked oxadiazoles as potential antimicrobial agents. Journal of Molecular Structure. 2024;1300:137260. doi: https://doi.org/10.1016/j.molstruc.2023.137260
- Zahra Z, Khan MR, Shah SA, Maryam S, Majid M, Younis T, et al. Vincetoxicum arnottianum ameliorate inflammation by suppressing oxidative stress and pro-inflammatory mediators in rat. Journal of Ethnopharmacology. 2020;252:112565. doi: https://doi.org/10.1016/j.jep.2020.112565
- Shamsi TN, Parveen R, Fatima S. Trypsin inhibitors demonstrate antioxidant activities, inhibit A549 cell proliferation, and increase activities of reactive oxygen species scavenging enzymes. Indian Journal of Pharmacology. 2017;49(2):155-160. doi: https://doi.org/10.4103/ijp.ijp_553_16
- Bhadane BS, Patil RH. Isolation, purification and characterization of antioxidative steroid derivative from methanolic extract of Carissa carandas (L.) leaves. Biocatalysis and Agricultural Biotechnology. 2017;10(2):216-223. doi: https://doi.org/10.1016/j.bcab.2017.03.012
- Galipalli S, Patel NK, Prasanna K, Bhutani KK. Activity-guided investigation ofCarissa carandas(L.) roots for anti-inflammatory constituents. Natural Product Research. 2014;29(17):1670-1672. doi: https://doi.org/10.1080/14786419.2014.989846
- Stærk D, Nezhad KB, Asili J, Emami SA, Ahi A, Sairafianpour M, et al. Phenanthroindolizidine alkaloids from Vincetoxicum pumilum. Biochemical Systematics and Ecology. 2005;33(9):957-960. doi: https://doi.org/10.1016/j.bse.2005.01.004
- Aksu D, Güzel Kara S. A Comparative Study on Some Biological Activities of Different Parts of Vincetoxicum hirundinaria. Hacettepe University Journal of the Faculty of Pharmacy. 2025;45(4):345-352. doi: https://doi.org/10.52794/hujpharm.1699849