RESEARCH ARTICLE
Avifaunal Diversity and Community Structure in the Forest Rehabilitation Zone of Mount Gede Pangrango National Park, Indonesia
Tropical Animals|Vol. 2, Issue 1, pp. 14-25 (2026)
CC BY 4.0-2026 Authors
Views
Downloads
Shares
Received
Apr 17, 2026Revised
May 29, 2026Accepted
Jun 28, 2026Published
Jun 30, 2026
Abstract
Habitat degradation in former production forests necessitates systematic rehabilitation to restore ecological functions and biodiversity. Monitoring avian communities serves as a vital bioindicator for evaluating the success of such restoration efforts. This study aimed to evaluate bird species diversity and community structure within the forest rehabilitation zone of Resort PTN Tapos, Mount Gede Pangrango National Park. Surveys were conducted in July 2023 using point count and MacKinnon list methods across observation points in the Cinakimun forest block. The study documented a relatively diverse bird community consisting of endemic and protected species, indicating that the rehabilitation area provides important habitat resources for avifauna. Ecological analyses demonstrated moderate species diversity, high distribution evenness, and low species dominance, reflecting a relatively stable bird community structure within the rehabilitated forest ecosystem. Vertical stratification observations further revealed that most bird activities were concentrated in the middle canopy layer, highlighting the ecological importance of vegetation structure in supporting bird habitat utilization. The findings indicate that the rehabilitation zone effectively supports a stable and diverse avian community, validating the success of current ecosystem recovery strategies in restoring vital wildlife habitats.
Keywords:
Introduction
Birds constitute one of the most widely distributed vertebrate groups, occupying nearly all vegetated ecosystems ranging from natural forests to anthropogenic landscapes. Among these, forest ecosystems particularly montane forests provide critical habitats due to their complex vertical structure and reduced anthropogenic disturbance, which enhance safety and resource availability for avifauna (1, 2). The presence and diversity of bird species within a habitat reflect ecological selection processes, where vegetation composition and structural complexity play pivotal roles in determining species richness and abundance. Habitats characterized by high vegetation diversity offer abundant food resources, nesting sites, and microhabitats, thereby supporting richer bird communities (3). Consequently, bird communities are widely recognized as reliable bioindicators for assessing ecosystem health and the success of habitat restoration efforts.
Despite their ecological importance, forest ecosystems are increasingly subjected to anthropogenic pressures, including land-use change and forest degradation, which threaten biodiversity and disrupt ecological functions. In Indonesia, many forest areas have undergone conversion to production forests, necessitating subsequent rehabilitation to restore ecological integrity. The Resort Pengelolaan Taman Nasional (PTN) Tapos, located within Taman Nasional Gunung Gede Pangrango (TNGGP), represents a critical case where former production forests are undergoing ecological rehabilitation to re-establish their conservation functions (4). The historical reliance on commercial monocultures, such as Pinus merkusii plantations, typically leaves a persistent structural and floristic legacy. This structural uniformity frequently acts as an ecological filter, constraining the recolonization of specialized montane avifauna by simplifying canopy layers and altering resource availability, which fundamentally shapes the baseline conditions for subsequent forest rehabilitation. Forest rehabilitation aims to restore vegetation cover, ecosystem processes. The rehabilitation zone in Resort PTN Tapos is characterized by montane forest habitat with diverse vegetation composition, including natural forest species and rehabilitated plant communities. The area is situated at a montane elevation range that supports relatively cool temperatures and high humidity, creating favorable environmental conditions for various bird species. In addition, the rehabilitation program in this area has been conducted for several years as part of ecosystem recovery efforts aimed at restoring vegetation structure, improving habitat quality, and increasing biodiversity conservation value. Forest rehabilitation aims to restore vegetation cover, ecosystem processes, and biodiversity, ultimately enhancing habitat suitability for wildlife, including birds (5). Previous studies in this area, such as in the Pasir Banteng block, recorded 32 bird species with moderate diversity (H’ = 2.76), relatively even species distribution (E = 0.79), and low dominance levels, indicating a recovering but not yet fully stable ecosystem (6). However, challenges remain in understanding how bird communities respond to ongoing rehabilitation processes, particularly in terms of community structure, diversity patterns, and conservation status.
Current approaches to evaluating ecosystem recovery often rely on vegetation-based indicators, which may not fully capture faunal responses and ecological interactions. Although avian diversity assessments have been conducted in certain parts of TNGGP, comprehensive studies focusing specifically on rehabilitation zones remain limited, creating a critical knowledge gap in understanding the trajectory of biodiversity recovery. Moreover, limited information is available regarding how bird communities adapt to rehabilitated montane habitats with varying vegetation structures and environmental conditions in Resort PTN Tapos. Addressing this gap is essential, as birds contribute significantly to ecological processes such as seed dispersal and pollination, thereby accelerating natural succession and enhancing forest resilience (7). Therefore, this study aims to analyze the diversity, community structure, and conservation status of bird species in the rehabilitation zone of Resort PTN Tapos, TNGGP. The research employs field-based observational methods combined with ecological indices, including species diversity, evenness, and dominance, to provide a comprehensive assessment of avian communities. The findings are expected to contribute to evidence-based conservation strategies and offer insights into the effectiveness of forest rehabilitation in restoring biodiversity and ecosystem functionality.
Methodology
Study Design and Study Area
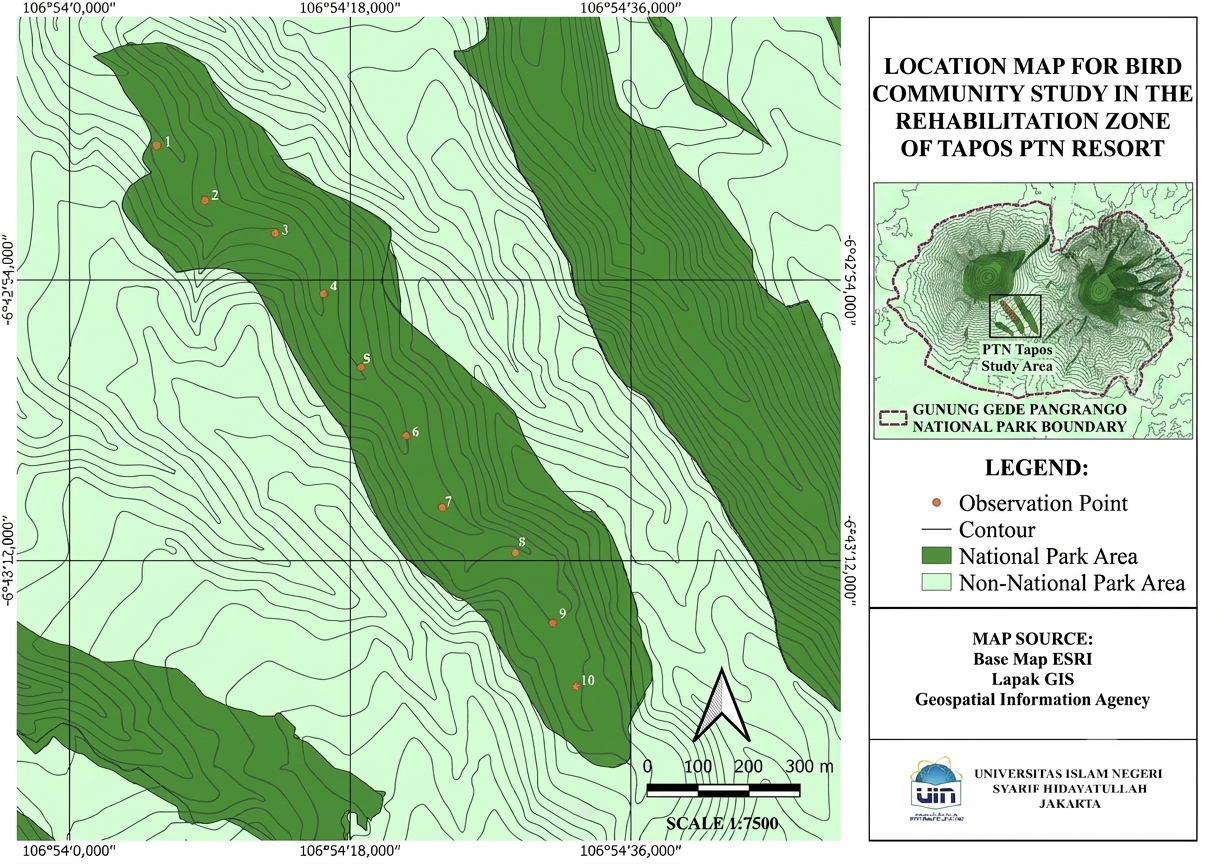
This study employed a quantitative ecological field survey designed to assess bird diversity, community structure, and habitat characteristics within a forest rehabilitation zone. The research was conducted in July 2023 in the rehabilitation area of Resort PTN Tapos, Taman Nasional Gunung Gede Pangrango (TNGGP), Indonesia. The study site is characterized by mixed secondary montane forest located within a hilly landscape adjacent to a river system. The spatial distribution of observation points is presented in Figure 1.
Data collection was conducted over a one-month period with eight repeated sampling events to improve detection probability and temporal representation. Observations were carried out twice daily during peak bird activity periods, specifically in the morning (05: 30–09: 00 WIB) and afternoon (15: 30–17: 00 WIB), based on the assumption that birds exhibit higher activity during these time intervals.
Bird Sampling Design and Procedures
Bird sampling was conducted using the point count method, which allows standardized estimation of bird presence and relative abundance in forest ecosystems. A total of 10 observation points were established along a linear transect, with a minimum distance of 150 m between points to avoid double counting of individuals. Each observation point had a fixed radius of 50 m and a standardized observation duration of 10 minutes, as illustrated in Figure 2.
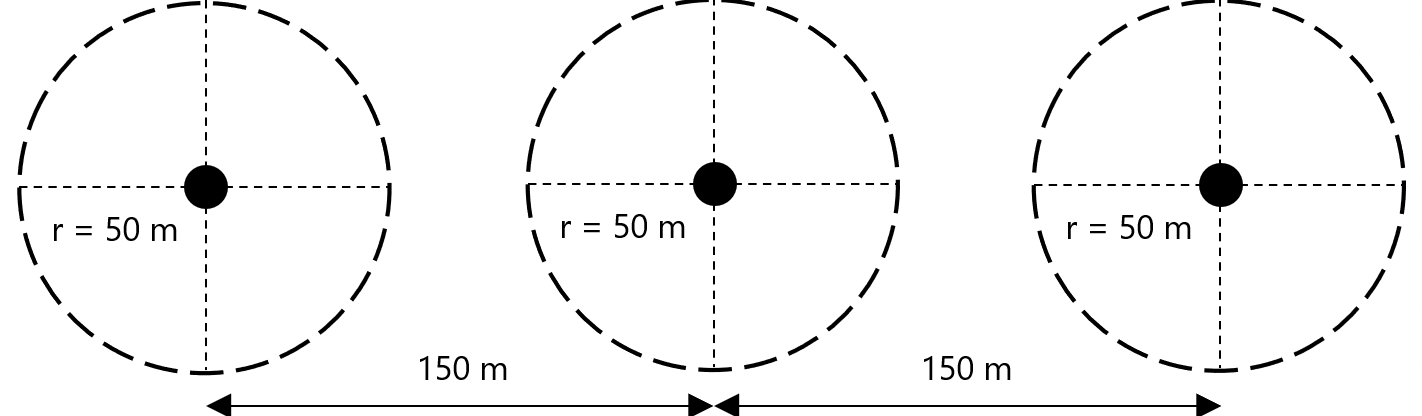
Sampling points were determined using purposive sampling based on habitat representativeness and areas with high bird encounter probability. At each point, all birds detected visually or acoustically were recorded. Observational variables included species identity, number of individuals, time of encounter, behavior, activity type, and vertical position within vegetation strata.
To complement the point count method, species richness was assessed using the MacKinnon list method, where bird species were sequentially recorded into lists containing a maximum of 20 species per list. Sampling continued until no additional species were recorded, indicating that the observed data sufficiently represented the bird community (8).
Bird Identification Techniques
Bird identification was conducted using both morphological and acoustic approaches. Morphological identification was based on key characteristics, including plumage coloration, pattern, body size, beak shape, and leg morphology, following standard field guides (8, 9). Acoustic identification was performed by recording bird vocalizations and cross-referencing them with validated databases such as Xeno-Canto, supported by automated recognition tools (BirdNET), to improve detection accuracy for cryptic and visually concealed species.
Taxonomic nomenclature, systematic ordering, and endemic designations strictly followed the most recent standardized Indonesian avifaunal checklist by Taufiqurrahman et al. (2022) (10). This ensures full compliance with contemporary taxonomic splits and updates relevant to Javan montane avifauna.
Measurement of Abiotic Factors
Abiotic environmental variables were measured at each observation point prior to bird surveys to characterize habitat conditions. Variables recorded included air temperature (°C), relative humidity (%), light intensity (lux), wind speed (m/s), and elevation (m above sea level). Measurements were conducted using a weather meter, luxmeter, and GPS device during both morning and afternoon sessions to capture temporal variation.
Vegetation Analysis
Vegetation structure was analyzed using a nested plot method to quantify habitat characteristics influencing bird communities. A total of five vegetation plots were established using a systematic-purposive approach to subsample the structural gradient across the 10 avifaunal observation points, with plots directly co-located at alternating bird sampling stations (Points 1, 3, 5, 7, and 9) at a spacing of 150 m between plots. These five plots serve as a representative macro-level characterization of the vegetation matrix, allowing structural metrics to be reliably correlated with broader avifaunal patterns along the transect. A total of five vegetation plots were established using purposive sampling to represent habitat variability, with a spacing of 150 m between plots. Each plot measured 20 × 20 m for the tree layer, with subplots of 10 × 10 m for poles and 5 × 5 m for saplings, as illustrated in Figure 3.
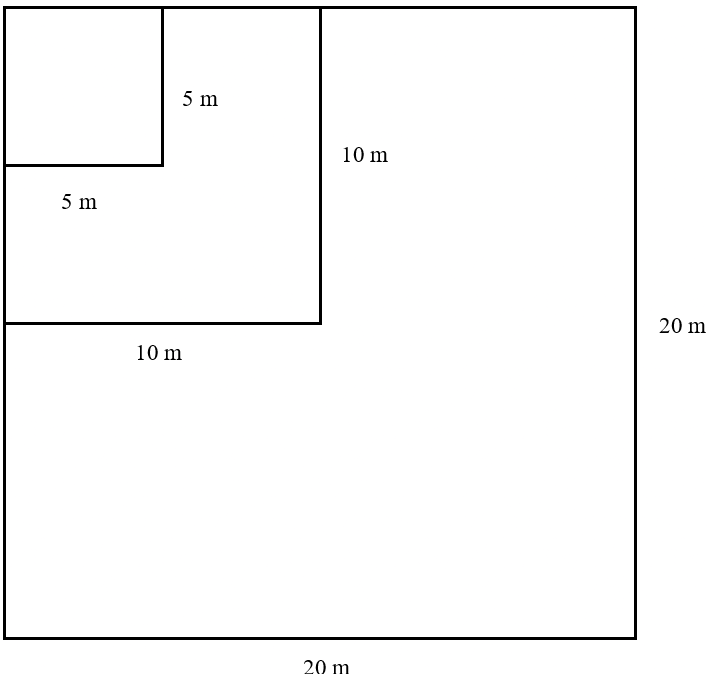
Vegetation was classified into growth stages, including saplings (height > 1.5 m and diameter < 10 cm), poles (diameter 10–20 cm), and mature trees (diameter > 20 cm). Data collected included species identity, number of individuals, tree height, height of the first branch, and diameter at breast height (DBH).
Data Analysis
Bird Community Analysis
Bird community structure was quantified using standard ecological indices. Species diversity was calculated using the Shannon–Wiener index (Equation 1), where Pi=NiNPi=NNi is the number of individuals of species i, and N is the total number of individuals. Diversity categories followed Odum (1993).
Species evenness was calculated using Equation 2, where S is the total number of species, with interpretation based on Krebs (1989) (11).
Species dominance was calculated Equation 3 following Odum (1993) (12). Species richness was calculated using the Margalef index (Equation 4) with interpretation based on Jorgensen et al. (2005) (13).
Vertical Distribution Analysis
The vertical distribution of bird species was analyzed descriptively based on vegetation strata classification (Table 1), following Indriyanto (14). This approach enabled the identification of habitat utilization patterns across forest canopy layers (15).
| No. | Vegetation Strata Category | Height Range (m) |
|---|---|---|
| 1. | Stratum A | > 30 |
| 2. | Stratum B | 20 - 30 |
| 3. | Stratum C | 4 - 20 |
| 4. | Stratum D | 1 - 4 |
| 5. | Stratum E | < 1 |
Species Composition and Conservation Status
Bird species composition was analyzed and presented in tables and graphs, including classification by family, feeding guild, and activity type. Conservation status was determined based on the IUCN Red List and national regulations (Permen LHK No. 106 Tahun 2018). Taxonomic nomenclature and endemicity classifications were systematically verified against Taufiqurrahman et al. (2022) (10) to maintain taxonomic uniformity throughout the study.
Vegetation Analysis
Vegetation data were analyzed using the Important Value Index (IVI), calculated as:
where FR represents relative frequency, KR relative density, and DR relative dominance. This index was used to determine the ecological importance of plant species within the study area:
Ethical Considerations
All data collection procedures were non-invasive and based on direct observation, ensuring minimal disturbance to wildlife. The study complied with conservation regulations within TNGGP, and all necessary research permits were obtained from the relevant authorities prior to fieldwork.
Results and Discussion
Habitat Conditions
The habitat characteristics at the study site were assessed through both abiotic and biotic components, including microclimatic variables and vegetation composition. The study area was located at an elevation range of 834–957 m above sea level (Appendix 2), representing a transitional zone between lowland and montane bird communities. Such altitudinal gradients are known to support mixed assemblages of bird species originating from both ecological zones (17), which is consistent with the species composition observed in this study (Table 2).
| No. | English Name | Latin Name | Family |
|---|---|---|---|
| 1 | Changeable Hawk-Eagle | Nisaetus cirrhatus | Accipitridae |
| 2 | Crested Serpent-Eagle | Spilornis cheela | Accipitridae |
| 3 | Collared Kingfisher | Todirhamphus chloris | Alcedinidae |
| 4 | Cave Swiftlet | Collocalia linchi | Apodidae |
| 5 | Yellow Bittern | Ixobrychus sinensis | Ardeidae |
| 6 | Bar-winged Prinia* | Prinia familiaris | Cisticolidae |
| 7 | Javan Tailorbird* | Orthotomus sepium | Cisticolidae |
| 8 | Spotted Dove | Spilopelia chinensis | Columbidae |
| 9 | Square-tailed Drongo-Cuckoo | Surniculus lugubris | Cuculidae |
| 10 | Banded Bay Cuckoo | Cacomantis sonneratii | Cuculidae |
| 11 | Rusty-breasted Cuckoo | Cacomantis sepulcralis | Cuculidae |
| 12 | Plaintive Cuckoo | Cacomantis merulinus | Cuculidae |
| 13 | Black Drongo | Dicrurus macrocercus | Dicruridae |
| 14 | Ashy Drongo | Dicrurus leucophaeus | Dicruridae |
| 15 | Javan Munia* | Lonchura leucogastroides | Estrildidae |
| 16 | Scaly-breasted Munia | Lonchura punctulata | Estrildidae |
| 17 | White-throated Kingfisher | Halcyon smyrnensis | Halcyonidae |
| 18 | Pacific Swallow | Hirundo tahitica | Hirundinidae |
| 19 | Flame-fronted Barbet* | Megalaima armillaris | Megalaimidae |
| 20 | White-crowned Forktail | Enicurus leschenaulti | Muscicapidae |
| 21 | Oriental Magpie-Robin | Copsychus saularis | Muscicapidae |
| 22 | Olive-backed Sunbird | Cinnyris jugularis | Nectariniidae |
| 23 | Little Spiderhunter | Arachnothera longirostra | Nectariniidae |
| 24 | Red Junglefowl | Gallus gallus | Phasianidae |
| 25 | Chestnut-bellied Partridge* | Arborophila javanica | Phasianidae |
| 26 | King Quail | Synoicus chinensis | Phasianidae |
| 27 | Fulvous-breasted Woodpecker | Dendrocopos macei | Picidae |
| 28 | Javan Banded Pitta* | Hydrornis guajana | Pittidae |
| 29 | Sooty-headed Bulbul | Pycnonotus aurigaster | Pycnonotidae |
| 30 | Common Moorhen | Gallinula chloropus | Rallidae |
| 31 | Pied Fantail | Rhipidura javanica | Rhipiduridae |
| 32 | Ashy Tailorbird | Orthotomus ruficeps | Sylviidae |
| 33 | Horsfield's Babbler | Malacocincla sepiaria | Timaliidae |
| 34 | Pygmy Wren-Babbler | Pnoepyga pusilla | Timaliidae |
| 35 | Rufous-fronted Laughingthrush* | Garrulax rufifrons | Timaliidae |
| 36 | Javan Cochoa* | Cochoa azurea | Turdidae |
| 37 | Javan White-eye | Zosterops melanurus | Zosteropidae |
| Note: *Endemic | |||
Microclimatic measurements indicated that mean air humidity was 84.23% in the morning and 77.65% in the afternoon (Appendix 3), values considered optimal for supporting avian activity (18). Humidity plays a critical role in regulating bird activity, as excessively high levels (> 90%) may constrain movement, whereas low humidity can increase dehydration risk and reduce activity levels (19, 20).
The average air temperature recorded was 21.42°C in the morning and 25.14°C in the afternoon (Appendix 3), which falls within the typical range for the Taman Nasional Gunung Gede Pangrango region. Temperature is a key ecological factor influencing bird distribution, as only species with suitable physiological adaptations can tolerate lower temperatures. Consequently, temperature variation contributes to shaping species composition and activity patterns within the habitat.
Wind speed measurements showed relatively low values, averaging 0.29 m/s in the morning and 0.38 m/s in the afternoon (Appendix 3), indicating stable atmospheric conditions. Such low wind velocities are unlikely to disrupt bird movement or behavior, as higher wind speeds are generally associated with reduced activity and increased risk during flight (21).
Light intensity ranged from 2.83 klux in the morning to 2.85 klux in the afternoon (Appendix 3), suggesting moderate illumination levels suitable for avian activity. Light availability influences daily behavioral patterns, with higher intensities often associated with reduced movement and increased resting behavior (22).
Vegetation analysis revealed a heterogeneous composition surrounding observation points, including Artocarpus heterophyllus, Calliandra calothyrsus, Coffea arabica, Nephelium lappaceum, Persea americana, Pinus merkusii, and Vernonia amygdalina (Appendix 1). Among these, Pinus merkusii was the most dominant species, with 31 individuals recorded, while Nephelium lappaceum was the least abundant (one individual). The dominance of Pinus merkusii is ecologically significant, as it provides perching and habitat structure for multiple bird species.
This dominance was further supported by the Importance Value Index (IVI), where Pinus merkusii exhibited the highest value (141.67%; Appendix 1), indicating its major ecological role in structuring the habitat. Rather than serving merely as a beneficial perching substrate, this non-native pine legacy carries profound ecological implications for forest rehabilitation. The structural and floristic uniformity characteristic of commercial pine legacy severely simplifies habitat architecture, often suppressing the regeneration of diverse native broadleaf flora (23). Consequently, this persistent monoculture framework acts as an active ecological filter that limits the recovery of specialized native montane avifauna, keeping the community characteristically reliant on edge-tolerant or generalist species during early-to-mid recovery phases. The high abundance and structural contribution of Pinus merkusii suggest that it acts as a primary determinant of habitat quality and resource availability for bird communities in the study area. However, the spatial asymmetry between the ten bird count stations and five vegetation plots presents a minor sampling limitation. While this sub-sampling design effectively captures macro-level environmental signals, it may underrepresent localized micro-topographical vegetation shifts—a constraint that highlights the need for fully paired, high-density spatial sampling in future long-term monitoring efforts.
Species Composition and Richness of Birds
A total of 37 bird species belonging to 24 families were recorded within the rehabilitation zone of Resort PTN Tapos. Among these, the family Cuculidae was the most represented, comprising four species: Surniculus lugubris, Cacomantis sonneratii, Cacomantis sepulcralis, and Cacomantis merulinus. The relatively high occurrence of Cuculidae is likely associated with the secondary forest habitat characterized by relatively open vegetation structure, which facilitates foraging efficiency and brood parasitism behavior (24). In addition, reduced competition and predation pressure may further contribute to their local abundance.
The complete list of recorded species is presented in Table 2, including several endemic taxa (marked with *), indicating the conservation importance of the study area. The presence of endemic species suggests that the habitat plays a critical role in supporting taxa with restricted geographic distributions and specialized ecological requirements.
Water-associated birds were represented by four species, namely Todirhamphus chloris, Halcyon smyrnensis, Ixobrychus sinensis, and Gallinula chloropus, each belonging to different families. Their occurrence indicates the availability of aquatic or semi-aquatic habitats that provide essential resources such as food, water, and breeding sites. Wetland-associated birds are known to be highly dependent on water availability and quality, making them reliable indicators of habitat condition (25).
Raptor species were represented by two taxa, Nisaetus cirrhatus and Spilornis cheela, which occupy the top trophic level within the ecosystem. The presence of these apex predators reflects a relatively balanced trophic structure, as raptors play a crucial role in regulating prey populations and maintaining ecosystem stability. Observations of soaring behavior further indicate active territory monitoring and foraging activity.
Ground-dwelling (terrestrial) bird species were also recorded, including Gallus gallus, Hydrornis guajana, Synoicus chinensis, and Arborophila javanica. These species are typically associated with forest floor habitats and are known to be sensitive to anthropogenic disturbance. Their presence suggests relatively low levels of human interference and adequate understorey vegetation that supports foraging and shelter requirements.
A total of eight endemic bird species were identified, including Prinia familiaris, Orthotomus sepium, Lonchura leucogastroides, Megalaima armillaris, Arborophila javanica, Hydrornis guajanus, Garrulax rufifrons, and Cochoa azurea. The occurrence of these species highlights the ecological significance of the area, as endemic taxa are often highly sensitive to habitat changes and can serve as important indicators for conservation prioritization (26).
Nectarivorous birds were represented by two species, Cinnyris jugularis and Arachnothera longirostra, indicating the presence of nectar-producing plants within the habitat. These species are considered ecological specialists, relying on specific floral resources, and their occurrence reflects habitat conditions that are still capable of supporting specialized ecological niches (27).
Woodpeckers were represented by a single species, Dendrocopos macei, which is typically associated with mature woody vegetation. The presence of this species suggests the availability of suitable tree substrates for foraging and nesting, particularly large or dead trees. This observation is consistent with the dominance of Pinus merkusii in the study area (Appendix 1), which provides structural habitat components required by woodpeckers (28).
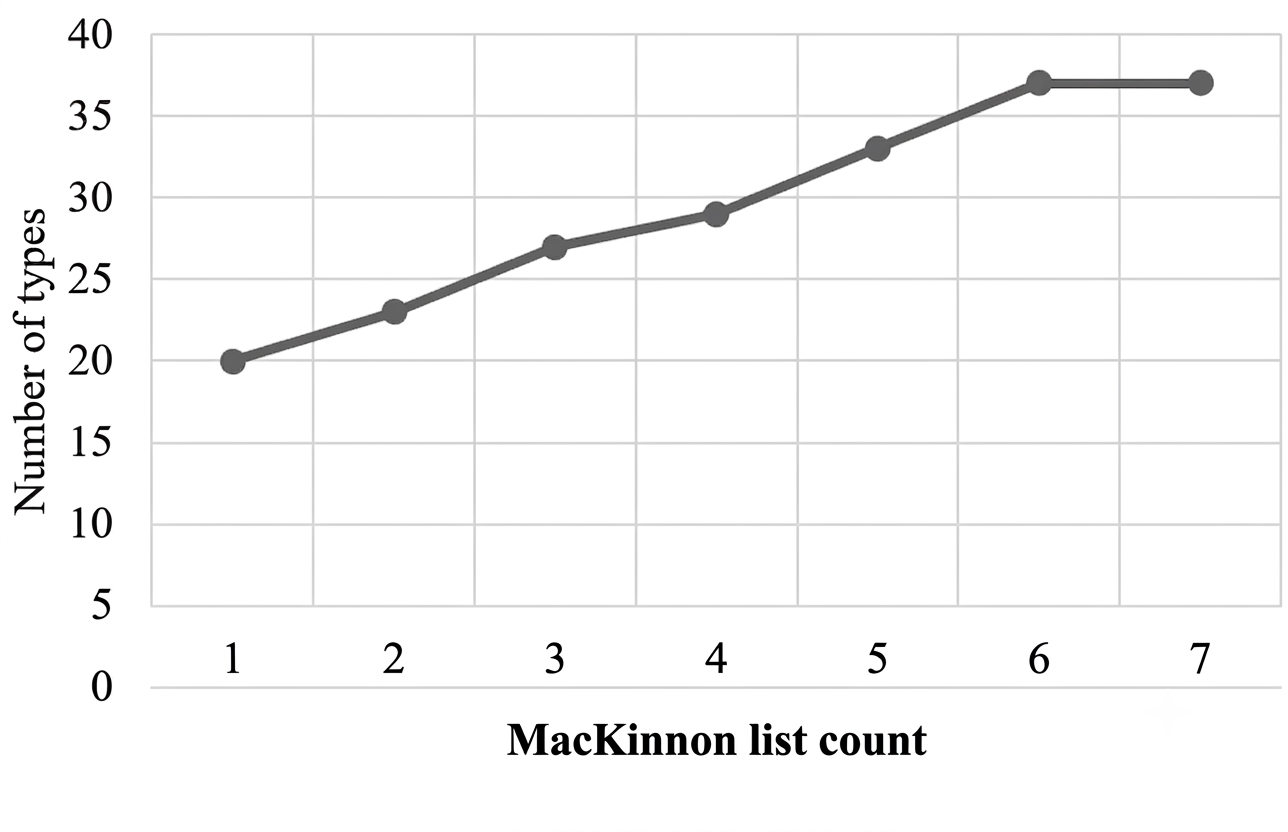
Species accumulation analysis based on the MacKinnon list method showed a progressive increase in species richness from the first to the fifth list, followed by a plateau between the sixth and seventh lists (Figure 4). This asymptotic pattern indicates that the sampling effort was sufficient to capture the majority of bird species present in the study area (8). Variations in species richness are generally influenced by factors such as resource availability, reproductive capacity, adaptive ability, and predation pressure within the ecosystem.
Distribution of Bird Species and Individuals
The spatial distribution of bird species varied across observation points, reflecting differences in habitat structure and environmental conditions. The highest species richness was recorded at point 7, with 19 species from 12 families (Figure 5A). This high occurrence is likely associated with favorable habitat characteristics, particularly the relatively open vegetation structure that provides diverse ecological resources such as food, perching sites, and shelter. Habitats with heterogeneous vegetation and sufficient resource availability are known to support higher bird diversity and distribution (29, 30). Open habitats, in particular, are frequently utilized by birds for foraging and thermoregulation activities, including sunbathing.
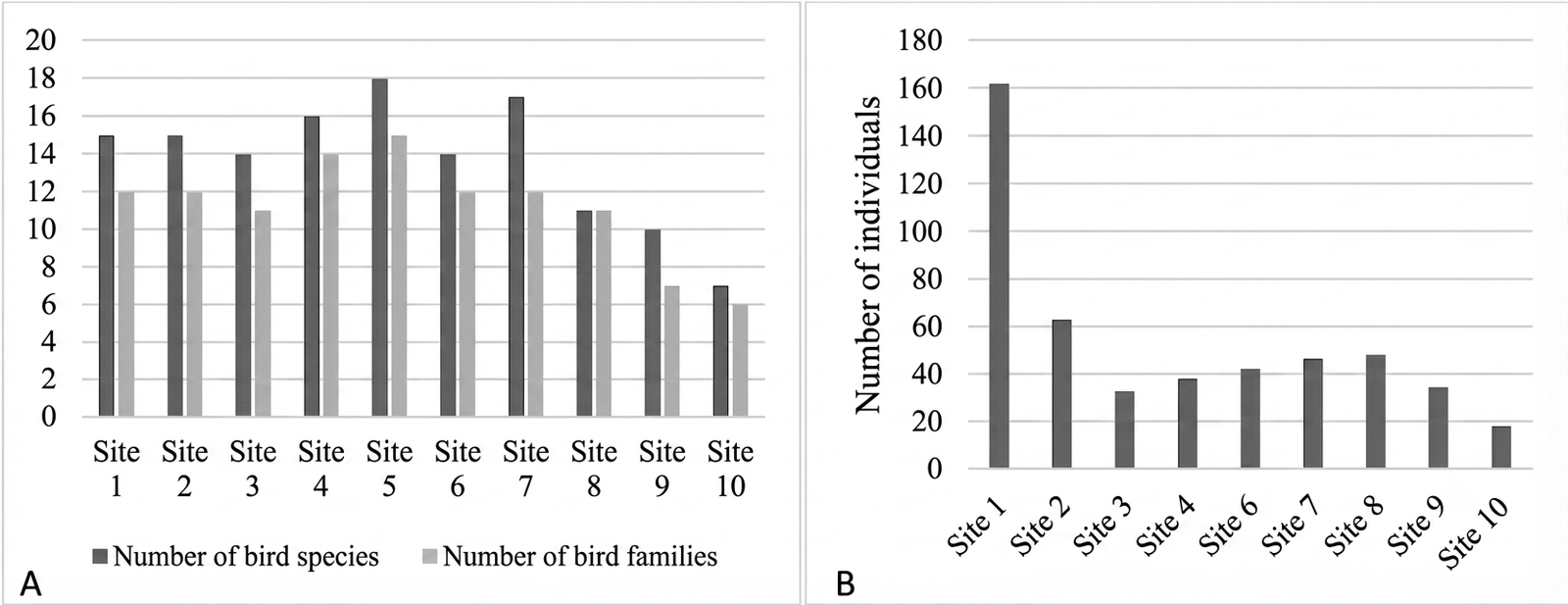
Conversely, the lowest species richness was observed at point 10, where only 7 species from 6 families were recorded (Figure 5A). This pattern can be attributed to denser vegetation structure and higher canopy closure, which may limit resource availability and restrict movement for certain bird species. In addition, point 10 represents the highest elevation among all observation sites (957 m a.s.l. ; Appendix 2), which may further contribute to reduced species richness. Increasing elevation is often associated with decreased vegetation diversity, ultimately influencing bird community composition (31).
The distribution of individual birds also showed distinct spatial variation. The highest abundance was recorded at point 1, with 162 individuals (31.76%) (Figure 5B). This site is characterized as a forest edge habitat with relatively open vegetation, conditions that are known to support higher bird abundance compared to interior forest habitats (32). Open and edge habitats tend to attract species that prefer accessible food resources and greater mobility. In this study, Lonchura leucogastroides and Lonchura punctulata were the most frequently observed species at point 1 (Appendix 4), consistent with their ecological preference for open grasslands and secondary vegetation, where they typically occur in pairs or small groups (8). This high concentration of open-country granivores at the forest boundary underscores intense edge effects and early-successional habitat characteristics. While the overall indices suggest a recovering system, the localized dominance of these generalist species serves as a clear indication of ongoing structural disturbance and habitat fragmentation at the rehabilitation zone’s perimeter, counterbalancing the narrative of an entirely uniform or stable interior forest ecosystem.
Bird Species Diversity
The analysis of bird species diversity at the study site indicated a moderate level of diversity (H′ = 3.00), suggesting that the ecosystem provides sufficient ecological support for a variety of bird species (Table 3). This condition reflects a relatively balanced ecosystem, as moderate diversity is typically associated with stable community structure and functional interactions among species. The presence of multiple ecological groups including waterbirds, terrestrial birds, woodpeckers, and raptors further supports this interpretation, indicating that the habitat accommodates a wide range of ecological niches.
| Parameter | Value | Criteria | Category |
|---|---|---|---|
| Diversity index (H′) | 3.00 | 1 < H′ ≤ 3 | Moderate |
| Evenness index (E) | 0.83 | E > 0.6 | High |
| Richness index (R) | 5.77 | R > 4 | Good |
| Dominance index (C) | 0.06 | C < 0.5 | No dominant species |
Vegetation composition is a key factor influencing bird diversity, as structurally and floristically diverse habitats tend to support higher species richness by providing varied food resources and microhabitats (33). This relationship is evident in the study area, where heterogeneous vegetation likely contributes to the observed diversity patterns.
The evenness index (E = 0.83) was categorized as high, indicating that individuals were relatively evenly distributed among species, with no single species numerically dominating the community. High evenness generally reflects stable resource availability and reduced interspecific competition (34). This is consistent with the low dominance index (C = 0.06), confirming the absence of dominant species and suggesting a well-balanced community structure (35).
The species richness index (R = 5.77) was classified as good, indicating that the study area supports a relatively high number of species. High richness values are commonly associated with habitats that offer diverse resources and environmental conditions suitable for multiple species. Such conditions reinforce the role of the study site as a suitable habitat for sustaining bird communities.
Comparative analysis with another study conducted within the same national park revealed differences in diversity values. Research by Kurnia & Mulawi (2023) reported a higher diversity index, with 50 species from 23 families. These differences may be attributed to variation in habitat conditions as well as sampling duration, as longer observation periods generally increase the likelihood of detecting more species (36). Temporal factors, including seasonal variation and species migration patterns, also influence bird diversity, potentially contributing to differences in observed richness and diversity across studies.
Bird Feeding Guilds
Bird species recorded in the study area were classified into six feeding guilds: frugivores, granivores, insectivores, carnivores, nectarivores, and omnivores (Figure 6). Among these, insectivores were the most dominant, comprising 18 species (49%), while nectarivores were the least represented, with only 2 species (5%). The presence of diverse feeding guilds indicates that the ecosystem is capable of providing a wide range of food resources required to sustain different trophic groups. Such trophic diversity also reflects the complexity of energy flow and ecological interactions within the habitat (37).
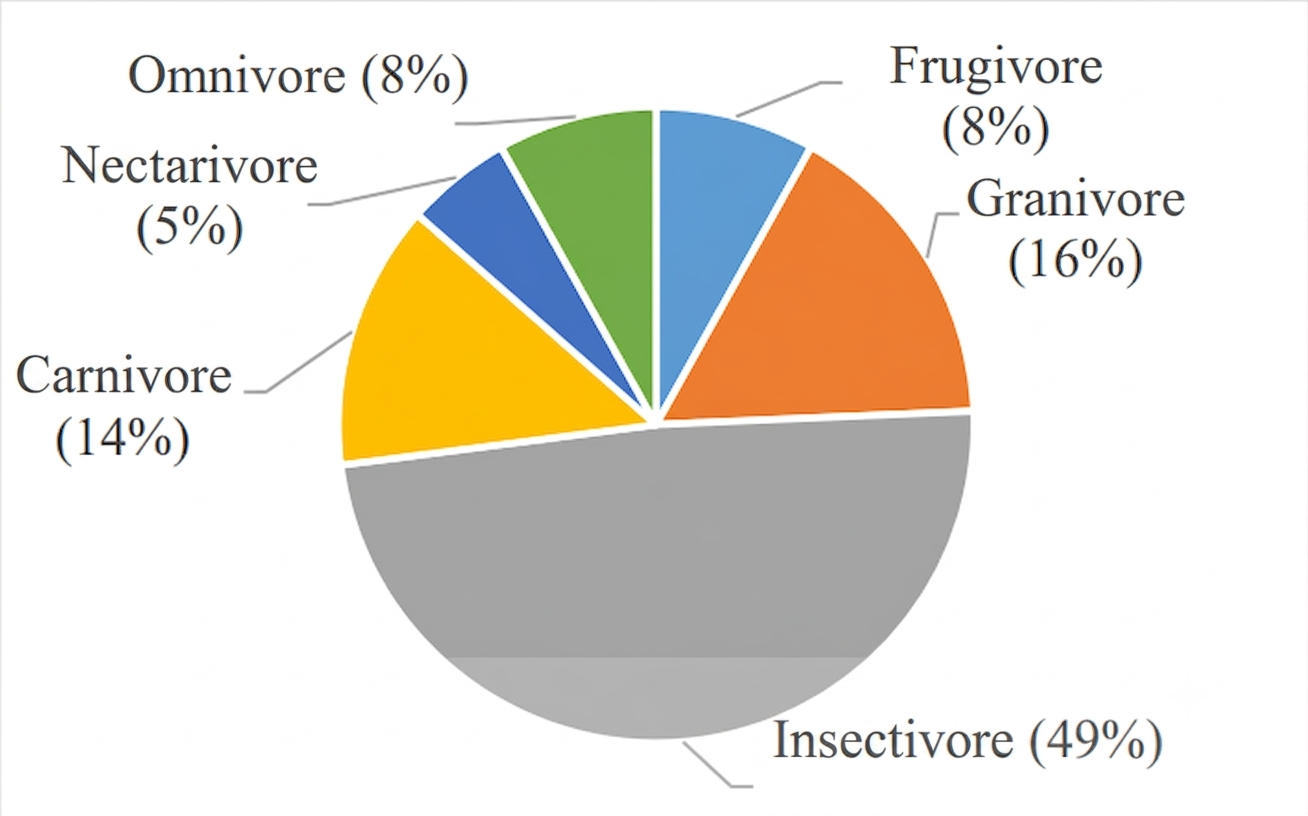
The dominance of insectivorous birds is a common pattern in forest ecosystems, where insects constitute a major and relatively тұрақ resource base. Globally, insectivores represent a substantial proportion of bird communities, often exceeding half of all species. This dominance is strongly associated with the continuous availability and diversity of insect prey, which supports a wide range of foraging strategies, from aerial hawking to bark gleaning (33). The adaptability of insectivorous birds to exploit various microhabitats further enhances their prevalence in such environments.
In contrast, nectarivorous birds were least encountered, represented only by Cinnyris jugularis and Arachnothera longirostra (Figure 6). Their low occurrence suggests limited availability of flowering plants producing nectar within the study area. Nectarivores are typically specialized feeders with narrow ecological niches, highly dependent on flowering phenology and plant availability (38). This interpretation is supported by the observation of only a single flowering plant species, Calliandra calothyrsus, during the study period. Additionally, the temporal nature of flowering cycles likely constrains nectar availability, thereby limiting the presence of nectar-feeding birds.
Vertical Distribution of Birds Across Vegetation Strata
Bird species in the study area utilized the full vertical profile of the forest, ranging from the upper canopy to the forest floor. The most frequently used vegetation stratum was Stratum C (4–20 m), which supported 16 species (25.56%), whereas Stratum B (20–30 m) was the least utilized, with only 2 species (Figure 7A; Table 1). Birds observed within these strata exhibited various activities, including flying, perching, foraging, vocalizing, and walking.
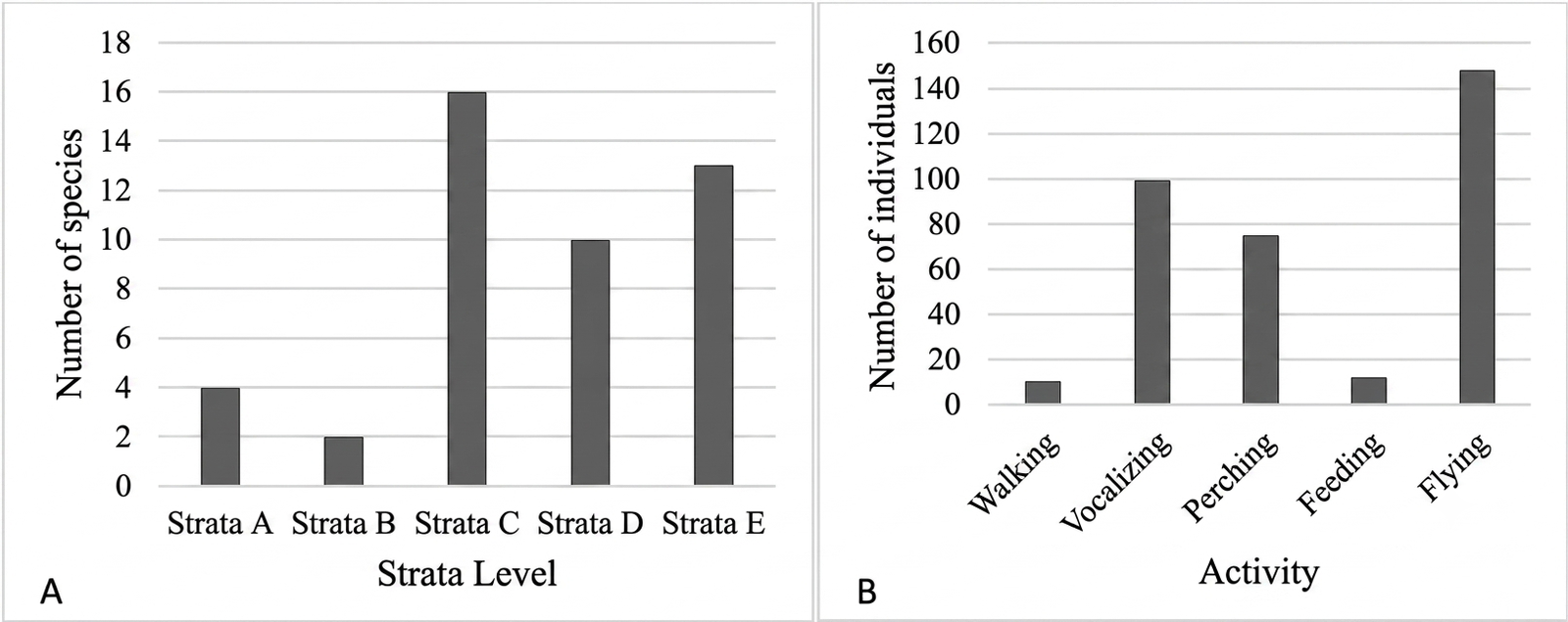
The high utilization of Stratum C indicates its ecological importance as a key habitat layer. This stratum likely provides optimal structural complexity and resource availability, supporting diverse bird activities. Mid-level vegetation offers suitable conditions for shelter from direct sunlight while maintaining distance from ground-level threats such as predators and human disturbance. In contrast, the low use of Stratum B may reflect the limited availability of trees within the 20–30 m height range, thereby restricting habitat suitability for many species (24). Nevertheless, certain species such as Collocalia linchi were observed in this stratum, likely due to favorable foraging conditions, including abundant aerial insects and fewer structural obstructions (39).
Some bird species were recorded utilizing multiple vegetation strata (Appendix 5), indicating flexibility in habitat use. This pattern reflects the differing ecological functions of each stratum and the need for birds to access various resources for feeding, resting, and nesting. Vertical stratification is therefore closely linked to resource partitioning and niche differentiation within bird communities (24).
Vertical distribution also corresponded with observed behavioral patterns. Flying was the most dominant activity, accounting for 148 individuals (42.9%) (Figure 7B). This may be associated with the availability of vegetation that facilitates movement and foraging across the habitat. In particular, some bird species rely heavily on aerial foraging strategies, especially insectivores that capture prey while in flight.
In contrast, walking was the least observed activity, with only 10 individuals (2.9%). This may be due to morphological constraints, as not all bird species are adapted for terrestrial locomotion. Additionally, limited understory vegetation may reduce opportunities for ground-based activities and increase exposure to predators. Ground-level vegetation such as shrubs and grasses typically provides important cover and foraging space for terrestrial birds, and its scarcity may further explain the low frequency of walking behavior.
Conservation Status of Bird Species
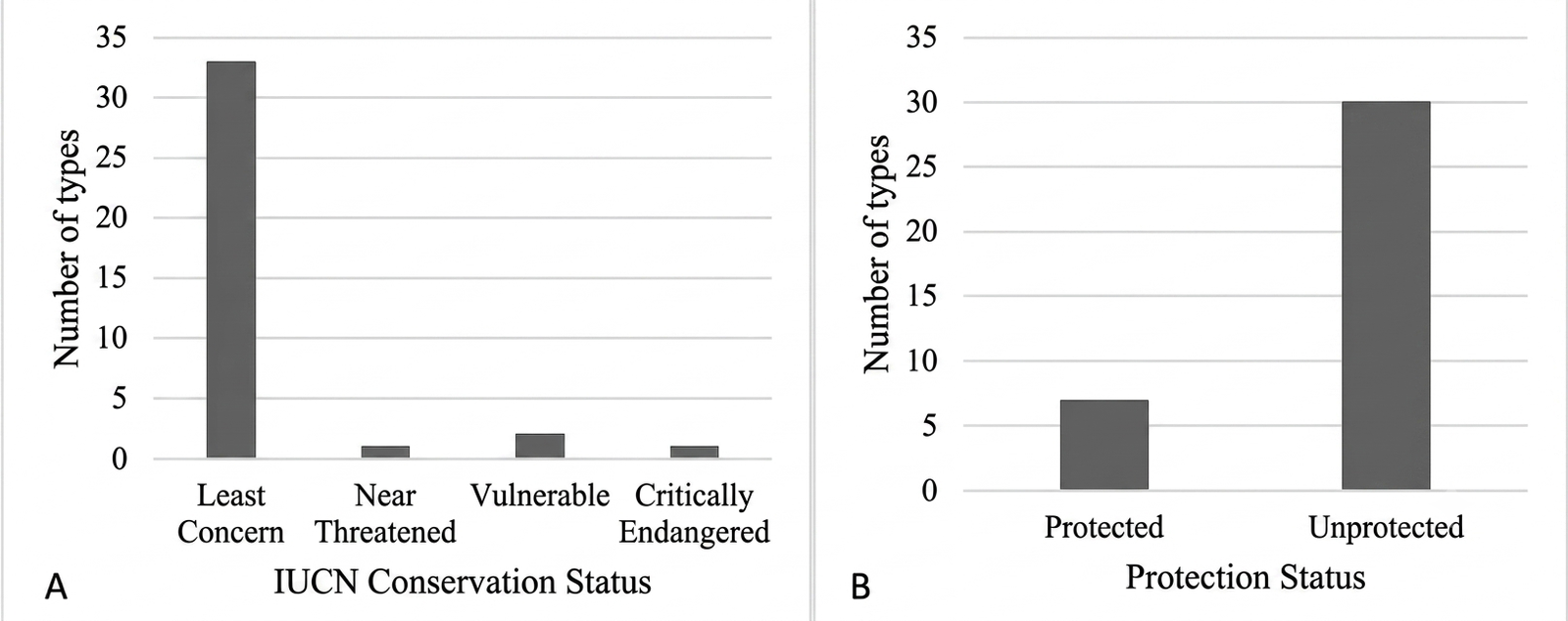
Based on the IUCN Red List, several bird species recorded in the study area fall into threatened categories (see Figure 8). Two species were classified as Vulnerable (VU), namely Cochoa azurea and Zosterops melanurus, while Prinia familiaris was categorized as Near Threatened (NT). Notably, Garrulax rufifrons was identified as Critically Endangered (CR), indicating a very high risk of extinction in the wild (Appendix 6). The detection of Garrulax rufifrons consisted of a small conspecific flock of three individuals, confirmed simultaneously via distinct acoustic vocalizations and clear visual contact. The flock was observed active within Stratum C (approximately 8–12 m above the ground), actively foraging and moving through the dense mid-canopy layer of mixed secondary vegetation. This specific microhabitat utilization highlights the value of complex middle-canopy structures in providing critical refugia for highly threatened endemic montane species.
The presence of Garrulax rufifrons (Figure 9) is particularly significant, as this species is known to face severe threats primarily due to intensive illegal trapping and its highly restricted distribution in montane forests of Java. The occurrence of a critically endangered species highlights the ecological importance of the study area and underscores the need for improved habitat management and conservation strategies. Birds are generally highly responsive to environmental changes, making them effective indicators of ecosystem condition (40).

In addition to global conservation status, national protection regulations (Permen LHK No. 106 Tahun 2018) identified seven protected species in the study area, including Nisaetus cirrhatus, Spilornis cheela, Megalaima armillaris, Hydrornis guajanus, Rhipidura javanica, Garrulax rufifrons, and Cochoa azurea (Appendix 6). These species are protected due to factors such as declining population trends, limited distribution, and endemicity. For example, Cochoa azurea is an endemic species with a restricted distribution in montane forests of Java, making it particularly vulnerable to habitat disturbance.
Interestingly, some species such as Nisaetus cirrhatus and Spilornis cheela are classified as Least Concern (LC) by the IUCN but are still protected under national regulations due to ongoing threats including deforestation, habitat degradation, and hunting pressure. The presence of protected raptors indicates a relatively intact trophic structure, as these species play a crucial role in maintaining ecological balance by regulating prey populations. Nisaetus cirrhatus and Spilornis cheela observed in the study area can be seen in Figure 10.

Conversely, Zosterops melanurus (VU) and Prinia familiaris (NT) are not currently protected under national regulations despite showing declining population trends. Their conservation status is largely driven by habitat loss and exploitation for the wildlife trade. This discrepancy between global and national conservation status suggests the need for reassessment and strengthened conservation measures. Effective strategies may include habitat protection, regular population monitoring, and control of illegal hunting to prevent further population declines (41). Zosterops melanurus (VU) and Prinia familiaris (NT) observed during the study can be seen in Figure 11.

Conclusion
The forest rehabilitation efforts at the PTN Tapos Resort have successfully fostered a stable and ecologically functional avian community, demonstrating an effective transition from a former production forest to a viable conservation zone. The documentation of 37 bird species, characterized by moderate biodiversity, high species evenness, and low dominance, indicates a balanced ecosystem with optimal resource partitioning. Furthermore, the presence of diverse trophic guilds, alongside eight Indonesian endemic and several globally threatened species, underscores the critical role of this recovering habitat as a vital wildlife refuge. Consequently, these findings empirically validate the current ecological restoration strategies and highlight the necessity for continuous habitat protection to ensure long-term avifaunal conservation.
Declarations
Conflict of Interest
The author declares no conflict of interest.
Data Availability
Data supporting the findings of this study are available from the corresponding author upon reasonable request.
Ethics Statement
Not applicable.
Funding Information
This work received no external funding.
Supplemental Material
The appendix is available as supplementary material at the following <a class="cursor-pointer" href="https://etflin.com/file/document/20260417015941_418830_a7757467.docx">link</a>.
References
- Kurniawan IS, Tapilouw FS, Hidayat T, Setiawan W. Keanekaragaman Aves di Kawasan Cagar Alam Pananjung Pangandaran. Jti. 2019;11(1):37-44. doi: https://doi.org/10.30599/jti.v11i1.393
- Hadiprayitno G, Mertha IG, Ilhamdi ML. Species Richness and Birds Conservation Priority in Mount Rinjani Areas, Lombok. J Bio Bio Edu. 2016;8(3):270. doi: https://doi.org/10.15294/biosaintifika.v8i3.5039
- Safanah NG. Keanekaragaman jenis burung di Taman Wisata Alam dan Cagar Alam Pananjung Pangandaran, Jawa Barat. Prosiding Seminar Nasional Masyarakat Biodiversitas Indonesia. 2017. doi: https://doi.org/10.13057/psnmbi/m030218
- Wahyuni AI, Khairiah A, Mulyawan B. Arboreal mammals inventory in Tapos area of gunung Gede Pangrango national park. BSc. 2022;6(1):72. doi: https://doi.org/10.24036/0202261114291-0-00
- ITTO. ITTO guidelines for the restoration, management and rehabilitation of degraded and secondary tropical forest. Japan: ITTO Policy Development Series 13; 2002.
- Kurnia I, Mulawi BA. Potensi Keanekaragaman Jenis Burung Untuk Birdwatching di Resort Situgunung dan Resort Cimungkad Taman Nasional Gunung Gede Pangrango. JB&P. 2023;10(1):14-24. doi: https://doi.org/10.29407/jbp.v10i1.19365
- Nurdin N, Nurlaila A, Kosasih D, Herlina N. Asosiasi Vegetasi Terhadap Komunitas Burung di Kampus I Universitas Kuningan. j. quagga. 2020;12(2):145. doi: https://doi.org/10.25134/quagga.v12i2.2672
- MacKinnon J, Phillipps K, Balen B Van. Burung-burung di Sumatera, Jawa, Bali dan Kalimantan: (termasuk Sabah, Sarawak dan Brunei Darussalam). Bogor: Puslitbang Biologi Lipi; 2010.
- Eaton JA, Balen B van, Brickle NW, Rheindt FE. Burung-burung Pulau Paparan Sunda dan Wallacea di Kepulauan Indonesia. Spain: Lynx Edicions; 2022.
- Taufiqurrahman I, Gusti Akbar P, Adi Purwanto A, Untung M. Panduan Lapangan Burung-Burung di Indonesia Seri 1: Sunda Besar. Birdpacker Indonesia; 2022.
- Krebs CJ. Ecological methodology. New York: Harper and Row Publ. Inc; 1989.
- Odum EP. Dasar-dasar Ekologi Terjemahan Tjahjono Samingan Edisi Ketiga. Yogyakarta: Gadjah Mada University Press; 1993.
- Jorgensen SE, Constanza R, Xu FL. Handbook of ecological indicators for assesment of ecosystem health. Boca Raton: CRC Press; 2005.
- Indriyanto. Ekologi Hutan. Jakarta: Bumi Aksara; 2006.
- Widodo W. Komparasi keragaman jenis burung-burung di taman nasional baluran dan alas purwo pada beberapa tipe habitat. Berkala Penelitian Hayati. 2009;14(2):113-124. doi: https://doi.org/10.23869/bphjbr.14.2.20091
- Sukmantoro W, Irham M, Novarino W, Hasudungan F, Kemp N, Muchtar M. Daftar Burung Indonesia no. 2. Bogor: Indonesian Ornithologists’ Union; 2007.
- Setiawan H. Kelimpahan dan Keanekaragaman Jenis Burung di Enclave Lindu Taman Nasional Lore Lindu. Biota : Jurnal Ilmiah Ilmu-Ilmu Hayati. 2014;18(2):100-109. doi: https://doi.org/10.24002/biota.v18i2.393
- Wulandari EY, Kuntjoro S. Keanekaragaman dan Kelimpahan Jenis Burung di Kawasan Cagar Alam Besowo Gadungan dan sekitarnya Kabupaten Kediri Jawa Timur. Jrba. 2019;1(1):18. doi: https://doi.org/10.26740/jrba.v1n1.p18-25
- Fajri MN, Kurnia I. Keanekaragaman Jenis Burung di Kecamatan Sukamakmur Kabupaten Bogor Provinsi Jawa Barat. Poltanesa. 2022;23(2):2092. doi: https://doi.org/10.51967/tanesa.v23i2.2092
- Xiong Y, Meng QS, Gao J, Tang XF, Zhang HF. Effects of relative humidity on animal health and welfare. Journal of Integrative Agriculture. 2017;16(8):1653-1658. doi: https://doi.org/10.1016/s2095-3119(16)61532-0
- Santillán V, Quitián M, Tinoco BA, Zárate E, Schleuning M, Böhning-Gaese K, et al. Spatio-temporal variation in bird assemblages is associated with fluctuations in temperature and precipitation along a tropical elevational gradient. PLoS ONE. 2018;13(5):e0196179. doi: https://doi.org/10.1371/journal.pone.0196179
- Rizki Syahputra MR, Ginantra IK, Raka Dalem AG. Aktivitas harian burung kuntul kecil (Egretta garzetta) di pulau serangan, BALI. Jbiounud. 2018;22(1):1. doi: https://doi.org/10.24843/jbiounud.2018.v22.i01.p01
- Destaranti N, Sulistyani S, Yani E. Struktur dan vegetasi tumbuhan bawah pada tegakan pinus di rph kalirajut dan rph baturraden banyumas. Scri. Biol. 2017;4(3):155. doi: https://doi.org/10.20884/1.sb.2017.4.3.407
- Dinanti RV, Winarni NL, Supriatna J. Vertical stratification of bird community in Cikepuh Wildlife Reserve, West Java, Indonesia. Biodiversitas. 2018;19(1):134-139. doi: https://doi.org/10.13057/biodiv/d190120
- Riefani MK, Soendjoto MA. Keragaman jenis burung air di kawasan Selat Sebuku Kotabaru, Kalimantan Selatan. In: Prosiding Seminar Nasional Pendidikan Biologi dan Biologi. 2015.
- Indrawan M, Primack RB, Supriatna J. Biologi Konservasi: Edisi Revisi. Jakarta: Yayasan Pustaka Obor Indonesia; 2007.
- Ulfah AN, Soendjoto MA, Peran SB, Wahyudi F. Keragaman spesies herba-liana dan kemiripan komunitasnya di area reklamasi pt adaro indonesia, provinsi kalimantan selatan, indonesia. Jss. 2020;3(3):432. doi: https://doi.org/10.20527/jss.v3i3.2176
- Kusmana C, Melyanti AR. Keragaman Komposisi Jenis dan Struktur Vegetasi pada Kawasan Hutan Lindung Dengan Pola PHBM di BKPH Tampomas, KPH Sumedang, Perum Perhutani Divisi Regional Jawa Barat dan Banten: Species Composition and Vegetation Structure of Protected Forest Area. J-Siltrop. 2017;8(2):123-129. doi: https://doi.org/10.29244/j-siltrop.8.2.123-129
- Purbawiyatna A, Kartodihardjo H, Alikodra HS, Prasetyo LB. Policy Analysis on Private Forest Management to Promote its Protectional Function. jpsl. 2012;2(1):1-10. doi: https://doi.org/10.19081/jpsl.2012.2.1.1
- Vikar A, Kartono AP, Mulyani YA. Komunitas burung pada ruang terbuka hijau di kota palu provinsi sulawesi tengah. Medkon. 2020;25(1):26-35. doi: https://doi.org/10.29244/medkon.25.1.26-35
- Seipalla B. Inventarisasi jenis burung pantai di kawasan pulau marsegu kabupaten seram bagian barat provinsi maluku. J.Hut.Trop. 2020;8(1):16. doi: https://doi.org/10.20527/jht.v8i1.8153
- Hasan N. Effect of relative humidity in an arid environment on dawn chorus. Journal of Entomology and Zoology Studies. 2017;562(51):562–4.
- Ekowati A, Setiyani AD, Haribowo DR, Hidayah K. Keanekaragaman jenis burung di kawasan telaga warna, desa tugu utara, cisarua, bogor. Al-Kauniyah J. Biol. 2016;9(2):3355. doi: https://doi.org/10.15408/kauniyah.v9i2.3355
- Lukmanul Hakim, Oekan S. Abdoellah, Parikesit, Withaningsih S. Impact of agricultural crop type and hunting on bird communities of two villages in Bandung, West Java, Indonesia. Biodiversitas. 2019;21(1). doi: https://doi.org/10.13057/biodiv/d210109
- Iswandaru D, Novriyanti N, Banuwa IS, Harianto SP. The Distribution of bird communities in University of Lampung, Indonesia. Biodiversitas. 2020;21(6). doi: https://doi.org/10.13057/biodiv/d210634
- Kurnia I, Mulawi BA. Potensi Keanekaragaman Jenis Burung Untuk Birdwatching di Resort Situgunung dan Resort Cimungkad Taman Nasional Gunung Gede Pangrango. JB&P. 2023;10(1):14-24. doi: https://doi.org/10.29407/jbp.v10i1.19365
- Muhammad GI, Mardiastuti A, Sunarminto T. Species and Feeding-Guild diversity of Avifauna at Gunung Pinang, Kramatwatu, Serang Regency, Banten. Medkon. 2018;23(2):178-186. doi: https://doi.org/10.29244/medkon.23.2.178-186
- Rofiq A, Harianto SP, Iswandaru D, Winarno GD. Guild pakan komunitas burung di kebun raya liwa kabupaten lampung barat. Jbl. 2021;4(2):195-206. doi: https://doi.org/10.29303/jbl.v4i2.753
- Manchi SS, Sankaran R. Foraging Habits and Habitat Use by Edible-nest and Glossy Swiftlets in the Andaman Islands, India. The Wilson Journal of Ornithology. 2010;122(2):259-272. doi: https://doi.org/10.1676/09-144.1
- Sanesi G, Padoa-Schioppa E, Lorusso L, Bottoni L, Lafortezza R. Avian Ecological Diversity as an Indicator of Urban Forest Functionality. Results from Two Case Studies in Northern and Southern Italy. isa. 2009;35(2):80-86. doi: https://doi.org/10.48044/jauf.2009.015
- Nainggolan FH, Dewi BS, Darmawan A. Status Konservasi Burung: Studi Kasus di Hutan Desa Cugung Kesatuan Pengelolaan Hutan Lindung Model Rajabasa Kecamatan Rajabasa Kabupaten Lampung Selatan (Bird Conservation Status: Case Study in Cugung Village Forest Kesatuan Pengelolaan Hutan Lindung Model Rajabasa Rajabasa Regency District South Lampung). Jsl. 2019;7(1):52-61. doi: https://doi.org/10.23960/jsl1752-61