RESEARCH ARTICLE
In Silico Study of Bioactive Compounds Fucoxanthin and Fucoidan from Sargassum echinocarpum as Anti-Cancer Agents Targeting Caspase-3 Protein
Aquatic Life Sciences|Vol. 1, Issue 2, pp. 45-52 (2025)
CC BY 4.0-2025 Authors
Views
Downloads
Shares
Received
Dec 25, 2024Revised
Jan 1, 2025Accepted
Jan 9, 2025Published
Jan 17, 2025
Abstract
Cancer is a complex disease characterized by disruptions in cell regulation mechanisms, leading to abnormal or uncontrolled cell growth and the ability to spread to other parts of the body. The caspase-3 protein plays a crucial role in the apoptotic mechanism and is a key target in cancer therapy. Bioactive compounds from the brown alga Sargassum, such as fucoidan and fucoxanthin, show significant potential as anticancer agents through mechanisms including apoptosis induction, cell cycle arrest, and metastasis inhibition. This study employed an in silico method to investigate the interaction between the natural ligands fucoidan and fucoxanthin and the target protein caspase-3, simulating their potential as anticancer drug candidates. The analysis revealed distinct interactions for each ligand with the target protein, as indicated by their binding affinity values. The interaction between fucoidan and caspase-3 resulted in a binding affinity value of ∆G -4.7 kcal/mol, which was lower in effectiveness compared to the interaction between fucoxanthin and caspase-3, with a binding affinity value of ∆G -7.7 kcal/mol. This suggests that fucoxanthin exhibits stronger anticancer activity through its interaction with caspase-3 compared to fucoidan. Meanwhile, toxicity tests showed that fucoidan has a lower toxicity profile than fucoxanthin, particularly in terms of LD₅₀ values and active/inactive toxicity parameters. These findings indicate that fucoxanthin has potential as anticancer drug candidates. Further research is needed to optimize their therapeutic efficacy and safety profiles.
Introduction
Cancer was first referred to as a tumor-like growth of tissue, a disease characterized by uncontrolled abnormal cell growth and the ability to spread to other parts of the body (1). If the spread of cancer cells, known as metastasis, is not stopped, it can lead to death (2). The causes of cancer involve various factors, both external such as exposure to tobacco, chemicals, radiation, and infection by certain organisms, and internal factors such as inherited genetic mutations, hormonal influences, immune system disorders, and random mutations (3). The carcinogenic mechanism involves a process in which carcinogens trigger changes in the genetic material (DNA) of cells or DNA damage. Carcinogens can damage DNA directly or interfere with DNA repair mechanisms, resulting in mutations (4). These mutations can activate oncogenes, inactivate tumor suppressor genes, or affect genes that regulate the cell cycle and apoptosis. The accumulation of such genetic changes leads to uncontrolled cell growth, resistance to cell death, and ultimately, malignant tumor formation (5).
An in-depth understanding of the molecular mechanisms underlying cancer development and control is key to finding more effective and safe therapies. One of the main focuses in cancer research is the apoptotic pathway, where caspase-3 acts as a key enzyme responsible for regulating programmed cell death. Cancer is referred to as a complex multifactorial disease with a high global mortality rate and a serious threat to human health. Moreover, cancer is the leading cause of death in developing countries and the second leading cause of death in developed countries (6). Caspase-3 is a key enzyme in the apoptotic (programmed cell death) pathway, and its role is crucial in cancer control. Apoptosis is the body's natural mechanism for eliminating damaged, old, or potentially cancerous cells (7). Caspases are a family of proteins that are highly homologous to the abnormal cell death-3 (CED-3) gene of C. elegans. Caspases can be divided into initiator caspases (caspase-2, -8, -9, -10), executioner caspases (caspase-3, -6, -7), and inflammatory caspases (caspase-1, -4, -5, -11) (8). Caspase-3 activation functions to break down target proteins, such as PARP (Poly ADP-Ribose Polymerase), which inhibits DNA repair and triggers cell death. It activates other enzymes that break down cell structures, thus bringing a controlled end to the cell's life cycle. Currently, chemotherapy is one of the most effective cancer treatments as it destroys cancer cells (9). However, this method has drawbacks, such as significant side effects and damage to healthy cells. Therefore, research into natural compounds is considered promising as an alternative or companion therapy that is potentially safer and more selective against cancer cells.
Sargassum echinocarpum is a type of brown algae that is known to have positive bioactivity for human health. The main components of brown algae consist of carbohydrates 33.9% - 70.0% (10) with 55% fucoidan (11), 40.5% alginate (12), and 34% laminaran (8). In addition, brown algae also contain 1.1%-26.8% protein, 0.6%-3.4% fat, 8.7%-41.2% minerals, and secondary metabolite components such as phenolic compounds and fucoidan (13). Fucoidan is a sulfated polysaccharide known to have biological activities, including antioxidant, anticancer, anti-inflammatory, and anticoagulant capabilities (14). Brown algae contain carotenoids such as fucosanthins, which are part of xanthophylls with high antioxidant activity (9). Fucoxanthin has a unique chemical structure containing nine conjugated double bonds, allenic bonds, epoxy, hydroxyl, carbonyl, and carboxyl groups in its molecule (15). Fucoxanthin exhibits a wide array of potential health benefits, including antioxidant activity, anti-inflammatory, anticancer, antiobesity, and antidiabetic effects, thus providing broad applications as a promising bioactive compound (12). The purpose of this study is to predict the bioactive potential of fucoidan and fucoxanthin in brown algae S. echinocarpum as anticancer drug candidates in silico. The in silico approach is used to analyze the detailed molecular interactions between fucoidan and fucoxanthin with target proteins such as caspase-3. This method is able to provide in-depth information before proceeding to in vitro or in vivo experiments.
Materials and Methods
Analysis of Bioactive Compounds and Target Proteins
The selection of bioactive compounds in brown algae S. echinocarpum was carried out through literature studies. The selected compounds as natural ligands are fucoidan (CID: 204) and fucosantin (CID: 5281239) whose molecular structures were taken from the web server https://pubchem.ncbi.nlm.nih.gov/. The target protein selected as modeling molecular docking analysis is caspase-3 (PDB ID: 2XYP) obtained from the web server https://www.rcsb.org/. Canonical SMILE of fucoidan and fucosantin compounds were used as drug candidate analysis using the web server https://www.way2drug.com/passonline/ (16).
Molecular Docking
Prediction of the potential of fucoidan and fucoxanthin compounds as anticancer drug candidates in silico was carried out using the PyRx 0.8 application. This method is to see how the interaction between natural ligands and target protein caspase-3. Parameters used in the molecular docking process include the value of Binding Affinity (ΔG), Hydrogen Bonding and Hydrophobic Interaction (16). In silico can be done by looking at the bonds formed between amino acids and what is the binding affinity value of each ligand to the target protein (17).
3D Molecular Docking Visualization
The molecular docking results were visualized in 3D using PyMOL software version 2.5.7. Analysis of binding interactions between fucoidan and fucoxanthin ligands with the target protein caspase-3 was performed with the help of the web server https://proteins.plus/ and Biovia Discovery Studio version 21.1.0. This visualization process allows more detailed identification of the type and location of molecular interactions formed (18).
Toxicity Test of Natural Ligands
Toxicity test analysis of natural ligands fucoidan and fucoxanthin as drug candidates was performed with the help of webserver https://tox.charite.de/protox3/index.php?site=home (19). The prediction of toxicity endpoints include acute toxicity, organ toxicity, toxicological endpoints, molecular initiating events, metabolism, adverse outcomes (Tox21) pathways and toxicity targets.
Results and Discussion
The 3D structure visualization of the natural ligand and target protein is shown in Figure 1. The target protein caspase-3 was chosen because of its directly responsible role in the mechanism of cell apoptosis in cancer (4). Apoptosis is a non-inflammatory form of cell death (PCD) mediated by the activation of apoptotic caspases and can occur through either the intrinsic pathway or the extrinsic pathway (20). The intrinsic pathway is activated by mitochondrial damage. Subsequently, cytochrome c is released into the cytoplasm from the mitochondria, combines with apoptotic protease activating factor-1 (Apaf-1) and caspase-9 precursors to form apoptosomes that activate caspase-9. The activated caspase-9 then cleaves and activates pro-caspase-3, which causes cell death by cleaving different cellular endogenous substrates (21). The extrinsic pathway is activated by cell surface death receptor signals, such as tumor necrosis factor-α (TNF-α) that binds to death receptors, then oligomerization of these receptors leads to recruitment and activation of caspase-8, which directly cleaves pro-caspase-3 to mediate apoptosis (7).
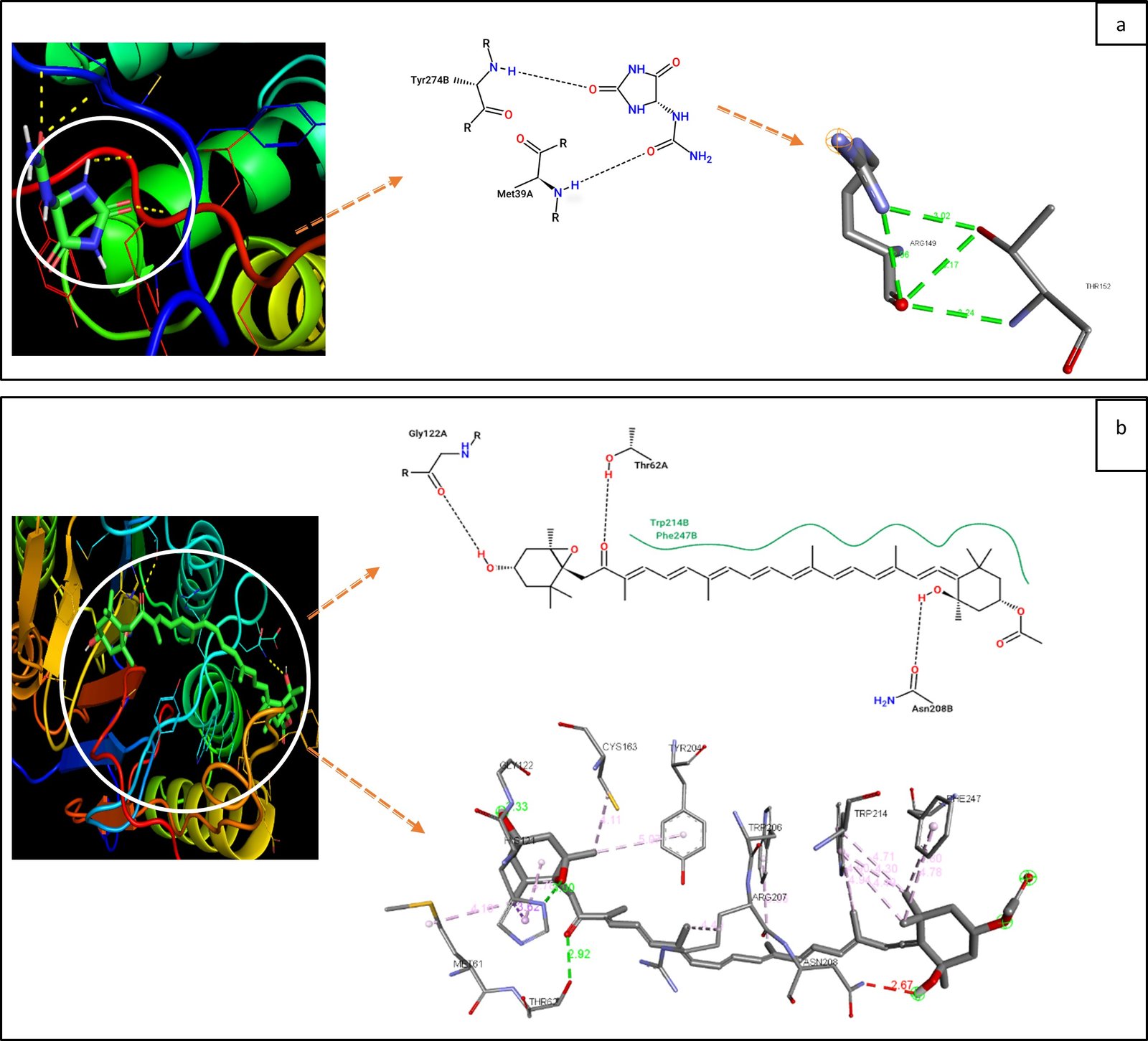
The binding affinity values obtained from the molecular docking process are presented in Table 1. The interaction between the natural ligand fucoidan and the target protein caspase-3 produces a binding affinity value of ∆G -4.7 Kcal/mol, which is higher when compared to the interaction between the natural ligand fucoxanthin and caspase-3, which has a binding affinity value of ∆G -7.7 Kcal/mol. This difference indicates that fucoxanthin exhibits a stronger interaction with caspase-3, suggesting a greater potential as an anticancer agent through the mechanism of interaction with this target protein (9). The stronger binding affinity of fucoxanthin is indicative of its ability to form more stable interactions with caspase-3, enhancing its bioactivity in inducing apoptosis in cancer cells. The structural characteristics of fucoxanthin, such as conjugated double bonds and reactive functional groups, contribute to its enhanced interaction with target proteins like caspase-3. These features allow fucoxanthin to establish more hydrogen bonds and hydrophobic interactions with key active residues on caspase-3, thereby stabilizing the enzyme's active conformation, which is a crucial step in the apoptotic pathway leading to cancer cell death (12). Furthermore, the unique chemical composition of fucoxanthin, including the presence of epoxide groups and polyene chains, contributes to its increased flexibility and stability within the active site of caspase-3 (see Figure 2). This structural advantage enhances its effectiveness in comparison to fucoidan, which, despite its biological activity, possesses a larger molecular structure with less flexibility, making it less effective in binding to and modulating the activity of caspase-3 (15). Previous studies have further supported these findings, demonstrating that fucoxanthin is not only more selective in inducing apoptosis in cancer cells but also exhibits broader health benefits, including antioxidant, anti-inflammatory, and anticancer properties (12). In contrast, although fucoidan has been reported to have various bioactive properties, its larger molecular size and structural rigidity limit its ability to interact effectively with the active sites of target proteins such as caspase-3. Due to its relatively weaker binding affinity, fucoidan may be less effective in triggering caspase-3 activation compared to fucoxanthin (15).
| Ligands | Binding affinity (Kcal/mol) | Amino Acids | |
| Hydrogen binding interactions | Hydrophobic interactions | ||
| Fucoidan | -4.7 | ASP40, MET39, TRY37 | TRY274 |
| Fucoxanthin | -7.7 | PHE247, ASN208, ARG207, TRP204, CYS163, GLY122, HIS121, MET61, THR62 | TRP214, TRP206 |
Fucoxanthin has shown potential as a cancer-preventive agent in various research models. The anticancer activity of fucoxanthin involves the induction of cell cycle arrest in the G1 and G2/M phases, which plays an important role in inhibiting cancer cell proliferation and apoptosis, while inhibiting the metastatic process, which is an important step in cancer therapy (22). Fucoxanthin was shown to also reduce cell survival and cell proliferation by inducing cell cycle arrest in several types of cancer cells, including human gastric adenocarcinoma cells SGC-7901 and BGC-823 (23), human lung cancer cells A549 and H1299 (24), and human bladder cancer cell line T24 (25). The mechanism of this proliferation inhibition involves increased expression of p21, a cyclin-dependent kinase (CDK) inhibitory protein, as well as decreased expression of CDK-2, CDK-4, cyclin D1, and cyclin E in T24 bladder cancer cells (26).
Previous research reported that fucoxanthin is able to induce apoptosis in SiHa human cervical cancer cells by increasing the expression of the pro-apoptotic protein Bax while decreasing the expression of the anti-apoptotic protein Bcl-2 (27). In addition, fucoxanthin is also known to increase the activity of caspase-3, a key enzyme in the apoptotic pathway.
Apoptosis induction by fucoxanthin is mediated through the inhibition of phosphatidylinositol-3-kinase (PI3K)/protein kinase B (Akt)/mechanistic target of rapamycin (mTOR) signaling pathway in cancer cells (28). These effects were also confirmed in vivo, where fucoxanthin administration inhibited polyp formation and increased cell death via an anoikis-like mechanism in the colonic mucosa of colorectal tumor-induced mice using azoxymethane and dextran sodium sulfate (29).
Other studies have revealed that fucoidan has anticancer potential through various mechanisms, including cell cycle arrest, inhibition of angiogenesis, induction of apoptosis, and activation of natural killer (NK) cells and macrophages (30). In addition, fucoidan is known to possess a remarkable array of biological activities, such as anti-inflammatory, antioxidant, anticoagulant, antithrombotic, antiviral, antiangiogenesis, and Helicobacter pylori infection-fighting effects (27). As a natural antioxidant, fucoidan is able to effectively counteract excess free radicals. In a study, low molecular weight fucoidan (LMWF) was processed into DF1, DF2, and DF3 fractions, all of which showed significant ability in scavenging superoxide anion radicals (31).
Based on the results of organ toxicity tests on fucoidan and fucoxanthin (see Table 2), there are significant differences in the toxicity levels of these two natural ligands. Fucoidan has an LD₅₀ value of 2600 mg/kg, so it is included in toxicity class 5 (very low toxicity) (14). In contrast, fucoxanthin has an LD₅₀ value of 130 mg/kg and belongs to toxicity class 3 (moderate toxicity) (15). These findings highlight that although fucoidan exhibits minimal toxicity, fucoxanthin, although effective in its bioactivity, should be considered with caution due to its moderate toxicity profile.
| Natural Ligand | Toxicity Test Results | |||
|---|---|---|---|---|
| LD50 | Toxicity Class | Inactive Toxicity | Active Toxicity | |
| Fucoidan | 2600mg/kg | 5 | Hepatotoxicity (0.53*) Nephrotoxicity (0.55*) Respiratory toxicity (0.54*) Cardiotoxicity (0.87*) Carcinogenicity (0.52*) Immunotoxicity (0.99*) Mutagenicity (0.64*) Cytotoxicity (0.69*) Nutritional toxicity (0.54*) | Neurotoxicity (0.57*) BBB-barrier (0.85*) Clinical toxicity (0.52*) |
| Fucoxanthin | 130mg/kg | 3 | Hepatotoxicity (0.81*) Neurotoxicity (0.8*) Cardiotoxicity (0.65*) Mutagenicity (0.5*) Cytotoxicity (0.70*) Ecotoxicity (0.67*) | Nephrotoxicity (0.69*) Respiratory toxicity (0.76*) Carcinogenicity (0.50*) Immunotoxicity (0.99*) BBB-barrier (0.59*) Clinical toxicity (0.61*) Nutritional toxicity (0.77*) |
| Note: *, Probability | ||||
In the inactive state, fucoidan showed a low risk of toxicity in most parameters, with probabilities below 0.6, except for immunotoxicity, which reached a very high value (0.99). Some other parameters, such as cardiotoxicity, also had probability values close to the safe limit (0.87), so they still need attention. On the other hand, fucoxanthin also has a low toxicity risk in the inactive state, but its immunotoxicity is as high as fucoidan (0.99), and other parameters, such as cardiotoxicity (0.65) and mutagenicity (0.50), still show a potential risk although it is relatively low.
In the active state, fucoidan showed some parameters with higher toxicity risks, such as neurotoxicity (0.57) and blood-brain barrier (BBB)-barrier (0.85), which require special attention. However, overall, the active toxicity of fucoidan remains lower than that of fucoxanthin. Fucoxanthin showed a higher risk of toxicity in some parameters, such as nutritional toxicity (0.77), neurotoxicity (0.69), and BBB-barrier (0.59), indicating that fucoxanthin has a greater potential risk in therapeutic use (12).
Overall, fucoidan has a safer toxicity profile than fucoxanthin, but the high immunotoxicity of both ligands remains an important concern. Therefore, further evaluation is needed, especially on high-probability toxicity parameters, to ensure the safety of both in medical or therapeutic applications.
According to Lin at al. (2020) added that the anti-cancer mechanism of fucoidan mainly includes the following four aspects (32). First, fucoidan can suppress the proliferation of cancer cells by inhibiting the normal mitosis of cancer cells and regulating the cell cycle. Second, fucoidan can activate the apoptotic signal of cancer cells, induce apoptosis through related pathways, and thus produce anticancer effects. The results of Kim at al. (2010) cultured HT-29 and HCT116, human colon cancer cells, with fucoidan extracted from Fucus vesiculosus. From the apoptosis detection results, fucoidan induced activation of caspase-3, -7, -8, -9, chromatin condensation and cleavage of poly (ADP-ribose) polymerase (PARP) (30). These data suggest that fucoidan can induce HT-29 and apoptosis of HVT116 cells through caspase-8 and -9-dependent pathways. Third, fucoidan can inhibit the formation of VEGF, thereby suppressing angiogenesis, cutting off the supply of tumor nutrients and oxygen, reducing tumor volume, and blocking the spread and displacement of cancer cells (33). Fourth, fucoidan can also activate the immune system, and then enhance the ability of natural killer cells and T cells to kill tumor cells. Fed mice that had been transplanted with NB4 acute promyelocytic leukemia cells with fucoidan, and it was found that fucoidan could effectively increase the killing activity of NK cells (34).
Conclusion
Fucoxanthin has more effective anticancer potential than fucoidan based on binding affinity values to the target protein caspase-3. However, its toxicity level was higher than fucoidan, which has a safer toxicity profile to human organs. Nonetheless, both ligands showed a significant risk of immunotoxicity, so further evaluation is needed to ensure their safety and effectiveness in medical applications.
Declarations
Conflict of Interest
The authors declare no conflicting interest.
Data Availability
The unpublished data is available upon request to the corresponding author.
Ethics Statement
Not applicable.
Funding Information
Not applicable.
References
- Brown JS, Amend SR, Austin RH, Gatenby RA, Hammarlund EU, Pienta KJ. Updating the Definition of Cancer. Mol Cancer Res. 2023;21(11):1142–7.
- Mueller T. What is the thalamus in zebrafish? Front Neurosci. 2012;6(MAY):1–14.
- Akter S. Pattern of Cancer and Its Risk Factor in Chattogram One Health Institute. 2022;
- de Almeida LC, Calil FA, Machado-Neto JA, Costa-Lotufo LV. DNA damaging agents and DNA repair: From carcinogenesis to cancer therapy. Vols. 252–253, Cancer Genetics. Elsevier Inc.; 2021. p. 6–24.
- Katerji M, Duerksen-Hughes PJ. Original Article DNA damage in cancer development: special implications in viral oncogenesis [Internet]. Vol. 11, Am J Cancer Res. 2021. Available from: www.ajcr.us/
- Teles MC, Cardoso SD, Oliveira RF. Social plasticity relies on different neuroplasticity mechanisms across the brain social decision-making network in zebrafish. Front Behav Neurosci. 2016;10(FEB).
- Jiang M, Qi L, Li L, Li Y. The caspase-3/GSDME signal pathway as a switch between apoptosis and pyroptosis in cancer. Vol. 6, Cell Death Discovery. Springer Nature; 2020.
- Kim OT, Jin ML, Lee DY, Jetter R. Characterization of the asiatic acid glucosyltransferase, UGT73AH1, involved in asiaticoside biosynthesis in centella asiatica (L.) Urban. Int J Mol Sci. 2017;18(12):1–11.
- Savira ADR, Amin MNG, Alamsjah MA. The effect of different type of solvents on the antioxidant activity of fucoxanthin extract from brown seaweed Sargassum duplicatum. In: IOP Conference Series: Earth and Environmental Science. IOP Publishing Ltd; 2021.
- Di Paolo C, Groh KJ, Zennegg M, Vermeirssen ELM, Murk AJ, Eggen RIL, et al. Early life exposure to PCB126 results in delayed mortality and growth impairment in the zebrafish larvae. Aquat Toxicol [Internet]. 2015;169:168–78. Available from: http://dx.doi.org/10.1016/j.aquatox.2015.10.014
- Palacios-filardo J, Mellor JR. Neuromodulation of hippocampal long-term synaptic plasticity. Curr Opin Neurobiol. 2019;54(Figure 2):37–43.
- Din NAS, Mohd Alayudin ‘Ain Sajda, Sofian-Seng NS, Rahman HA, Mohd Razali NS, Lim SJ, et al. Brown Algae as Functional Food Source of Fucoxanthin: A Review. Vol. 11, Foods. MDPI; 2022.
- Zhang Y, Hawboldt K, MacQuarrie S. Extraction of bioactive compounds from beach-cast brown algae: a review on accelerated solvent extraction and subcritical water extraction. Vol. 2, RSC Sustainability. Royal Society of Chemistry; 2024. p. 2069–91.
- Kashyap B, Saikia K, Samanta SK, Thakur D, Banerjee SK, Borah JC, et al. Kaempferol 3-O-rutinoside from Antidesma acidum Retz. Stimulates glucose uptake through SIRT1 induction followed by GLUT4 translocation in skeletal muscle L6 cells. J Ethnopharmacol. 2023 Jan 30;301.
- Saravana PS, Cho YJ, Park YB, Woo HC, Chun BS. Structural, antioxidant, and emulsifying activities of fucoidan from Saccharina japonica using pressurized liquid extraction. Carbohydr Polym. 2016 Nov 20;153:518–25.
- Wardani DWKK, Ali M, Khotimah H, Nurdiana, Riawan W, Primihastuti D, et al. The effect of Centella asiatica to the vascular endothelial growth factor and vascular endothelial growth factor receptor-2 on the rotenone induced zebrafish larvae (Danio rerio) stunting model. GSC Biol Pharm Sci. 2018;5(2):088–95.
- Hao Y, Huang J, Ma Y, Chen W, Fan Q, Sun X, et al. Asiatic acid inhibits proliferation, migration and induces apoptosis by regulating Pdcd4 via the PI3K/Akt/mTOR/p70S6K signaling pathway in human colon carcinoma cells. Oncol Lett. 2018;15(6):8223–30.
- Schöning-Stierand K, Diedrich K, Ehrt C, Flachsenberg F, Graef J, Sieg J, et al. ProteinsPlus: a comprehensive collection of web-based molecular modeling tools. Nucleic Acids Res. 2022 Jul 5;50(W1):W611–5.
- Banerjee P, Kemmler E, Dunkel M, Preissner R. ProTox 3.0: A webserver for the prediction of toxicity of chemicals. Nucleic Acids Res. 2024 Jul 5;52(W1):W513–20.
- Rogers C, Fernandes-Alnemri T, Mayes L, Alnemri D, Cingolani G, Alnemri ES. Cleavage of DFNA5 by caspase-3 during apoptosis mediates progression to secondary necrotic/pyroptotic cell death. Nat Commun. 2017 Jan 3;8.
- Ahmed FF, Abd El-Hafeez AA, Abbas SH, Abdelhamid D, Abdel-Aziz M. New 1,2,4-triazole-Chalcone hybrids induce Caspase-3 dependent apoptosis in A549 human lung adenocarcinoma cells. Eur J Med Chem. 2018 May 10;151:705–22.
- Mills CC, Kolb EA, Sampson VB. Development of chemotherapy with cell-cycle inhibitors for adult and pediatric cancer therapy. Vol. 78, Cancer Research. American Association for Cancer Research Inc.; 2018. p. 320–5.
- Jing Y, Wang G, Ge Y, Xu M, Tang S, Gong Z. AA-PMe, a novel asiatic acid derivative, induces apoptosis and suppresses proliferation, migration, and invasion of gastric cancer cells. Onco Targets Ther. 2016;9:1605–21.
- Mei CH, Zhou SC, Zhu L, Ming JX, Zeng FD, Xu R. Antitumor effects of laminaria extract fucoxanthin on lung cancer. Mar Drugs. 2017 Feb 15;15(2).
- Wang L, Zeng Y, Liu Y, Hu X, Li S, Wang Y, et al. Fucoxanthin induces growth arrest and apoptosis in human bladder cancer T24 cells by up-regulation of p21 and down-regulation of mortalin. Acta Biochim Biophys Sin (Shanghai). 2014 Oct 1;46(10):877–84.
- Bae M, Kim MB, Park YK, Lee JY. Health benefits of fucoxanthin in the prevention of chronic diseases. Vol. 1865, Biochimica et Biophysica Acta - Molecular and Cell Biology of Lipids. Elsevier B.V.; 2020.
- Jin Y, Qiu S, Shao N, Zheng J. Fucoxanthin and tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) synergistically promotes apoptosis of human cervical cancer cells by targeting PI3K/Akt/NF-kB signaling pathway. Med Sci Monit. 2018 Jan 1;24:11–8.
- Dou H, Yu P yang, Liu Y qi, Zhu Y, Li F cheng, Wang Y yu, et al. Recent advances in caspase-3, breast cancer, and traditional Chinese medicine: a review. Vol. 36, Journal of Chemotherapy. Taylor and Francis Ltd.; 2024. p. 370–88.
- Terasaki M, Iida T, Kikuchi F, Tamura K, Endo T, Kuramitsu Y, et al. Fucoxanthin potentiates anoikis in colon mucosa and prevents carcinogenesis in AOM/DSS model mice. J Nutr Biochem. 2019 Feb 1;64:198–205.
- Nagamine T, Kadena K, Tomori M, Nakajima K, Iha M. Activation of NK cells in male cancer survivors by fucoidan extracted from cladosiphon okamuranus. Mol Clin Oncol. 2020 Jan 1;12(1):81–8.
- van Weelden G, Bobi M, Okła K, van Weelden WJ, Romano A, Pijnenborg JMA. Fucoidan structure and activity in relation to anti-cancer mechanisms. Mar Drugs. 2019 Jan 7;17(1).
- Lin Y, Qi X, Liu H, Xue K, Xu S, Tian Z. The anti-cancer effects of fucoidan: A review of both in vivo and in vitro investigations. Vol. 20, Cancer Cell International. BioMed Central Ltd.; 2020.
- Ireson C, Orr S, Jones DJL, Verschoyle R, Lim CK, Luo JL, et al. Characterization of metabolites of the chemopreventive agent curcumin in human and rat hepatocytes and in the rat in vivo, and evaluation of their ability to inhibit phorbol ester-induced prostaglandin E2production. Cancer Res. 2001;61(3).
- Atashrazm F, Lowenthal RM, Woods GM, Holloway AF, Karpiniec SS, Dickinson JL. Fucoidan Suppresses the Growth of Human Acute Promyelocytic Leukemia Cells In Vitro and In Vivo. J Cell Physiol. 2016 Mar 1;231(3):688–97.