
RESEARCH ARTICLE
Evaluation of Pork-Specific DNA Primers for Halal Authentication in Processed Meats
Halal Science|Vol. 1, Issue 1, pp. 25-30 (2025)
Views
Downloads
Shares
Received
May 25, 2025Revised
Jun 16, 2025Accepted
Jun 30, 2025Published
Jun 30, 2025
Abstract
Keywords:
Introduction
The global halal food industry, valued at over USD 2 trillion, continues to grow as consumers seek products that meet religious and safety standards. However, recurring cases of undeclared pork contamination have raised concerns about the reliability of halal labeling (1, 2). In Islamic dietary law, the concept of halalan thayyiban, meaning permissible and wholesome, forms a fundamental requirement. While many food products claim to meet these standards, increasing reports of adulteration, particularly with pork (Sus scrofa) derivatives, have raised significant concerns regarding the integrity of processed meat products in the market (3).
The adulteration of halal-labeled processed meats with pork is not only deceptive but poses ethical, religious, and health-related challenges for Muslim consumers (4, 5). Several studies have documented the intentional mixing of pork in various meat products such as sausages, meatballs, and corned beef, primarily for economic gain (6). This issue is exacerbated by the physical similarity between processed pork and other meats, making visual detection unreliable. Hence, scientific verification methods are urgently needed to ensure compliance with halal standards. Current detection methods primarily rely on protein-based assays such as immunoassays and electrophoresis. While useful, these techniques suffer from significant limitations (7). Proteins degrade or denature under high processing temperatures and pressures, leading to false negatives in processed food matrices (8). Consequently, DNA-based methods, particularly Polymerase Chain Reaction (PCR), have emerged as more reliable alternatives due to DNA's higher stability under extreme processing conditions and its species-specific resolution (9).
PCR techniques have proven effective in amplifying target DNA sequences, including mitochondrial markers like Cytochrome B, for species identification (10, 11). However, the effectiveness of PCR is highly dependent on the specificity and sensitivity of the primers used (12). Despite several studies employing various pork-specific primers, inconsistencies in amplification efficiency and sensitivity across different food matrices suggest that not all primers are equally robust (13). Moreover, there is limited comparative data assessing the relative sensitivity of multiple pork-specific primers under uniform experimental conditions. To address this gap, the present study systematically evaluates the sensitivity of five pork-specific DNA primers, ND5, Cytochrome B (1), Cytochrome B (2), PPA8, and Pork, in detecting pork DNA within processed meat products. This approach offers a critical assessment of primer performance and identifies the most reliable molecular markers for pork authentication. The novelty of this study lies in its head-to-head comparison of multiple primers targeting different mitochondrial genes within the same experimental context. By doing so, the study not only determines which primers offer the highest detection sensitivity but also provides a foundational toolset for halal authentication protocols in the food industry. The aim of this study is to determine the most sensitive pork-specific DNA primers for the detection of Sus scrofa DNA in processed meat products using PCR analysis. To this end, laboratory-based assays were conducted on corned pork and beef meatball samples to evaluate the amplification performance of each primer.
Experimental Section
Study Design and Sample Collection
This study was designed as an exploratory laboratory-based analysis to evaluate the sensitivity of five pork-specific DNA primers in detecting Sus scrofa DNA within processed meat products. The selected food matrices included commercially available corned pork and beef meatballs, representing positive and negative sample groups respectively. Beef meatball was chosen as the negative control sample due to its high-fat, processed matrix characteristics, which resemble those of corned pork. This ensures a comparable food matrix for assessing potential cross-reactivity and evaluating PCR specificity under similar processing conditions. Both sample types were obtained from retail outlets across Surabaya, Indonesia. Upon acquisition, samples were immediately stored at 4 °C during transport and transferred to a −20 °C freezer upon arrival at the laboratory to preserve DNA integrity prior to extraction.
DNA Extraction Protocol
Genomic DNA was extracted using the QIAamp DNA Mini Kit (QIAGEN, Germany), with procedural optimization for high-fat processed food matrices. The QIAamp DNA Mini Kit utilized in this study operates on a silica membrane-based solid phase extraction principle, enabling effective removal of PCR inhibitors commonly found in processed meat matrices. This method was selected for its reliability in obtaining high-quality DNA suitable for downstream PCR analysis. Approximately 25 mg of homogenized sample material was transferred into a sterile 1.5 mL microcentrifuge tube. To this, 180 µL of buffer ATL and 20 µL of Proteinase K were added, and the mixture was vortexed for 15 s to ensure homogenization. Samples were then incubated overnight at 60 °C (12–16 h) to ensure complete lysis and protein digestion.
Following lysis, 200 µL of buffer AL was added to each sample and vortexed briefly, followed by an additional incubation step at 70 °C for 10 min to enhance nucleic acid release. An equal volume (200 µL) of 96% ethanol was subsequently added to the lysate and mixed by vortexing. The entire mixture was then transferred into a QIAamp Mini spin column inserted into a 2 mL collection tube and centrifuged at 8,000 rpm for 1 minute. The flow-through was discarded, and the column was washed sequentially with 500 µL of Buffer AW1 and Buffer AW2, centrifuged at 8,000 rpm and 14,000 rpm respectively. The final elution was carried out using 100 µL of Buffer AE, which was added to the membrane and incubated at room temperature for 5 min before a final centrifugation at 8,000 rpm for 1 minute. The purified DNA was stored at −20 °C until analysis. DNA concentration and purity were assessed using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific), and integrity was verified by agarose gel electrophoresis.
Primer Selection and Design
Five pork-specific primers were selected based on their previously validated specificity and application in food DNA testing. These included ND5, Cytochrome B (1), Cytochrome B (2), PPA8, and a pork-specific primer known as Pork (F2/R1). Each primer targets a distinct mitochondrial DNA locus with varying annealing temperatures and amplicon lengths (see Table 1). The ND5 primer targets the NADH dehydrogenase subunit 5 gene; Cytochrome B primers amplify different regions within the mitochondrial cytochrome b gene; PPA8 targets the mtATP8 region; and the Pork (F2/R1) primer pair amplifies a 130 bp region also within the cyt b gene. Primers were synthesized commercially (Integrated DNA Technologies) and reconstituted to a working concentration of 10 µM. Details of primer sequences, target genes, product sizes, and annealing temperatures were verified through NCBI primer-blast alignment.
| Primer Name | Forward Primer Tm (°C) | Reverse Primer Tm (°C) |
|---|---|---|
| ND5 | 55.6 | 55.3 |
| Cytochrome B (1) | 47.1 | 50.7 |
| Cytochrome B (2) | 57.5 | 58.5 |
| Pork (F2/R1) | 46.9 | 50.0 |
| PPA8 | 56.4 | 54.3 |
| Note: Tm = Melting temperature calculated for primer-template hybridization optimization. | ||
Polymerase Chain Reaction (PCR) Conditions
Conventional PCR amplification was conducted in a total volume of 25 µL using a T100 Thermal Cycler (Bio-Rad, USA). Each reaction mixture contained 12.5 µL of 2× PCR Master Mix (Intron Biotechnology, South Korea), 1 µL each of forward and reverse primers (10 µM), 2 µL of DNA template (approximately 50–100 ng), and 8.5 µL of nuclease-free water. The thermal cycling profile began with an initial denaturation at 95 °C for 5 min, followed by 35 amplification cycles consisting of denaturation at 95 °C for 30 s, primer-specific annealing for 30 s (50–58 °C, depending on the primer), and extension at 72 °C for 30 s. A final extension step was performed at 72 °C for 5 min. All reactions included negative controls (no template) and beef-only DNA controls to monitor for contamination or nonspecific amplification.
Electrophoresis and Visualization
PCR amplicons were analyzed via agarose gel electrophoresis using 1.5% agarose gels prepared in 1× TBE buffer. Gels were stained with RedSafe™ nucleic acid staining solution and loaded with 5 µL of PCR product mixed with 1 µL of 6× loading dye. A 100 bp DNA ladder (GeneDireX) served as the molecular size marker. Electrophoresis was conducted at 100 V for 45 min, and DNA bands were visualized using a UV transilluminator (UVP BioDoc-It Imaging System). Sensitivity was determined based on the presence and clarity of DNA bands at the expected size, and primer performance was evaluated by comparing the signal strength across replicate reactions.
Data Analysis and Sensitivity Assessment
The sensitivity of each primer was defined by its ability to reliably amplify pork DNA at low concentrations and across sample replicates. Positive detection was indicated by the appearance of clear, sharp DNA bands corresponding to the expected amplicon size. Visual comparison of band intensity was used to semi-quantitatively rank primer performance, while cross-reactivity was assessed using beef meatball samples as negative controls. Experimental repeatability and consistency were ensured by conducting each PCR assay in triplicate.
Results
DNA Quality and Yield from Processed Meat Samples
High-quality genomic DNA was successfully extracted from both corned pork and beef meatball samples using a modified silica column-based protocol. Visual inspection of agarose gels confirmed the presence of intact genomic DNA with minimal degradation across all samples. The extracted DNA displayed consistent purity, with A260/A280 ratios ranging between accepted values, indicating low protein contamination. DNA concentrations were sufficient for downstream PCR applications, with no observable inhibition in negative controls, suggesting the effectiveness of the extraction method in processed meat matrices. These baseline quality metrics are summarized in Table 2.
| Sample | Purity (A260/A280) | Concentration (ng/µL) |
|---|---|---|
| Pig (corned) | 1.64 | 26.5 |
| Cow (meatball) | 1.87 | 31.6 |
PCR Amplification Across Five Primers
PCR assays were performed using five pork-specific primers to evaluate their ability to amplify pork DNA in corned meat samples while ensuring specificity against beef-derived matrices. Table 2 presents the amplification performance of each primer across the two sample types. Clear amplification bands corresponding to the expected sizes were observed for three of the five primers tested. The primers targeting mtATP8 and the two cytochrome b variants, PPA8, Pork (F2/R1), and Cytochrome B (2), consistently yielded strong, well-defined bands in corned pork samples, while no amplification occurred in beef-only samples. In contrast, ND5 and Cytochrome B (1) primers produced either weak or no amplification, suggesting suboptimal sensitivity under the tested conditions. The primer sequence amplification can be seen in Table 3.
| No. | Primer name | DNA sequence | Product size (bp) |
|---|---|---|---|
| 1 | ND5 | F: 5’-AGC TGC ACT ACA AGC AAT CC-3’ | 227 |
| R: 5’-ATG CGT TTG AGT GGG TTA GG-3’ | |||
| 2 | Cytochrome B (1) | F: 5’-ATG ACC AAC ATC CGA AAA TC-3’ | 1140 |
| R: 5’-TCA TTT TAA TAG GTT GTT TTC G-3’ | |||
| 3 | Cytochrome B (2) | F: 5’-CCA TCC AAC ATC TCA GCA TGA TGA AA-3’ | 359 |
| R: 5’-GCC CCT CAG AAT GAT ATT TGT CCT CA-3’ | |||
| 4 | Pork | F: 5’-CTT GCA AAT CCT AAC AGG CCT G-3’ | 130 |
| R: 5’-CGT TTG CAT GTA GAT AGC GAA TAA C-3’ | |||
| 5 | PPA8 | F: 5’-ATC TAC ATG AAT CAT TAC AAT TAC-3’ | 126 |
| R: 5’-CTA TGT TTT TGA GTT TTG AGT TCA-3’ |
Gel Electrophoresis Visualization
Representative gel images depicting the amplification outcomes are shown in Figure 1. DNA bands were present only in corned pork samples for the three more sensitive primers, confirming target specificity and lack of cross-reactivity. The electrophoresis patterns demonstrated consistent reproducibility across triplicate reactions. Notably, samples amplified using PPA8 and Pork (F2/R1) primers produced bands with higher visual intensity compared to Cytochrome B (2), suggesting stronger amplification efficiency.
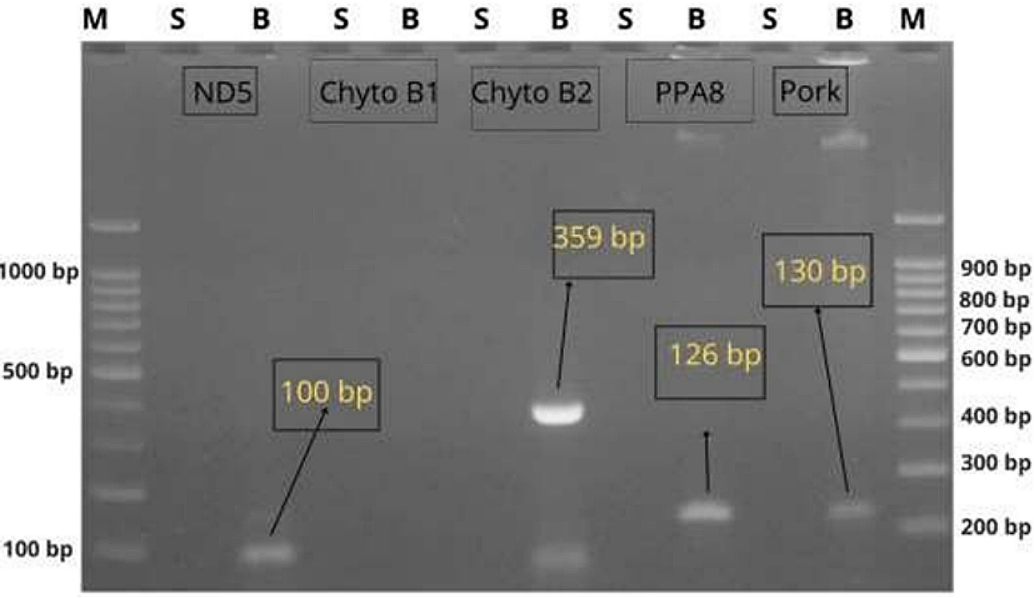
Comparative Sensitivity Analysis
Sensitivity comparison among the primers revealed that PPA8 and Pork (F2/R1) primers demonstrated the highest capacity to detect pork DNA in processed matrices. These findings can be seen in Table 4, where only three primers produced reliable signals, indicating that primer-target sequence compatibility and annealing temperatures play crucial roles in amplification performance. The absence of bands in beef samples across all primer sets further supports the specificity of the positive amplifications.
Discussion
The present study aimed to evaluate and compare the sensitivity of five pork-specific DNA primers in detecting Sus scrofa DNA within processed meat products, with the goal of identifying robust molecular markers for halal food authentication. The results demonstrated that only three of the five primers—PPA8, Pork (F2/R1), and Cytochrome B (2)—were capable of consistently amplifying pork DNA under the conditions tested. This finding is in line with previous reports suggesting that not all primers targeting mitochondrial genes are equally effective when applied to complex food matrices subjected to thermal and chemical processing (14).
Table 4 shows a clear distinction in performance among the primers. The ND5 and Cytochrome B (1) primers failed to produce detectable bands in the pork samples, indicating either insufficient sensitivity or non-optimal annealing characteristics. These outcomes are consistent with prior findings by Anggita (2019), who reported that ND5, while specific, had reduced performance at low DNA concentrations or when amplifying degraded DNA, as typically found in processed foods (15). In contrast, primers PPA8 and Pork (F2/R1) yielded strong, sharp amplicons at the expected sizes, confirming their high sensitivity and suitability for detection of trace pork DNA, even after food processing. These findings are corroborated by Tanabe et al. (2007), who identified the F2/R1 primer pair as a reliable marker in multiple food matrices with minimal false positives (16).
| Primer | Sample | Amplified | Observed band size (bp) | Expected band size (bp) |
|---|---|---|---|---|
| ND5 | Corned pork | Yes | 100 | 227 |
| Cytochrome B (1) | Corned pork | Yes | – | 1, 140 |
| Cytochrome B (2) | Corned pork | Yes | 359 | 359 |
| PPA8 | Corned pork | Yes | 126 | 126 |
| Pork (F2/R1) | Corned pork | Yes | 130 | 130 |
| ND5 | Beef meatball | No | – | – |
| Cytochrome B (1) | Beef meatball | No | – | – |
| Cytochrome B (2) | Beef meatball | No | – | – |
| PPA8 | Beef meatball | No | – | – |
| Pork (F2/R1) | Beef meatball | No | – | – |
The amplification visualized in Figure 1 supports this conclusion. Clear bands were observed exclusively in pork samples for PPA8, Pork (F2/R1), and Cytochrome B (2), with no cross-reactivity seen in beef meatball controls. The stronger band intensities associated with PPA8 and Pork suggest a higher amplification efficiency, likely due to better primer-target binding affinity and compatibility with the annealing temperature used. Notably, Cytochrome B (2) also amplified pork DNA successfully, albeit with lower visual intensity, pointing to its acceptable but comparatively moderate sensitivity.
This performance variance among primers may be attributed to differences in target gene copy number, sequence stability, and fragment size (17-19). Mitochondrial DNA is favored in species identification due to its abundance in cells and resistance to degradation (20). However, certain mitochondrial regions may be more stable or accessible post-processing. For instance, the mtATP8 gene targeted by PPA8 may be more conserved or less prone to fragmentation under thermal stress, which could explain its superior amplification results in this study. These findings align with observations by Yoshida et al. (2009), who reported high amplification success from mtATP8 across a range of mammalian tissues (21).
The inability of ND5 and Cytochrome B (1) to detect pork DNA highlights a common issue in primer design: theoretical specificity does not always translate into functional performance in real-world matrices (22). These primers may still be effective under optimized conditions or in fresh tissue but are evidently less suited for processed meat authentication without further refinement. This underlines the need for rigorous empirical validation of primers before routine diagnostic or regulatory use, especially in halal authentication where false negatives could mislead consumers and regulators. Importantly, no amplification was detected in the beef meatball samples across all five primers, reinforcing the specificity of the primers that produced positive results. This absence of cross-reactivity is critical for halal compliance testing, where sensitivity must be balanced with species-level specificity. As shown in Table 4, the successful primers discriminated effectively between pork and beef DNA, confirming their practical utility.
Overall, this study contributes valuable comparative data to the field of halal food authentication by providing a side-by-side evaluation of commonly used pork-specific primers. The findings underscore the importance of primer choice in PCR-based meat species identification and suggest that PPA8 and Pork (F2/R1) are currently among the most reliable for detecting Sus scrofa DNA in processed foods. Future studies could build on these results by evaluating primer performance across broader sample types, including mixed matrices and highly degraded products, or by integrating quantitative PCR to assess detection thresholds.
Although the conventional PCR approach applied here demonstrated clear species-specificity, one limitation of this study is the use of only two sample types, corned pork and beef meatballs, which may not fully represent the complexity of commercial meat products. Additionally, band intensity was evaluated visually, which, while informative, lacks the precision of quantitative PCR. Future work should include a broader sample matrix and employ real-time PCR to determine detection limits and enable more robust quantification. To summarize, PPA8 and Pork (F2/R1) primers consistently produced strong, specific amplification of pork DNA without cross-reactivity in beef samples, indicating their superior suitability for routine halal food verification.
Conclusion
This study demonstrated that among the five pork-specific DNA primers evaluated, PPA8, Pork (F2/R1), and Cytochrome B (2) showed the highest sensitivity and specificity for detecting Sus scrofa DNA in processed meat products using conventional PCR. These primers consistently amplified pork DNA without cross-reactivity to beef, confirming their potential as reliable tools for halal food authentication. Future research should explore the performance of these primers across a wider range of processed food matrices, under various processing conditions, and incorporate quantitative PCR approaches to determine detection thresholds and enhance analytical precision.
Declarations
Conflict of Interest
The authors declare no conflicting interest.
Data Availability
The unpublished data is available upon request to the corresponding author.
Funding Information
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
References
- Alfie Syarifah Y, Putri EBP. Factors Related to Consumer Preferences of Halal Certified Restaurants (Case Study in Fast Food Restaurant X). J Halal Res Policy, Ind. 2022 Jul 31;1(1):1–7.
- Usman H, Chairy C, Projo NWK. Between awareness of halal food products and awareness of halal-certified food products. J Islam Mark. 2023 Feb 10;14(3):851–870.
- Kang TS, Tanaka T. Comparison of quantitative methods based on SYBR Green real-time qPCR to estimate pork meat adulteration in processed beef products. Food Chem. 2018 Dec;269:549–558.
- Hamdan H, Issa ZM, Abu N, Jusoff K. Purchasing Decisions among Muslim Consumers of Processed Halal Food Products. J Food Prod Mark. 2013 Jan;19(1):54–61.
- Aisyah M. The Influence of Religious Behavior on Consumers’ Intention to Purchase Halal-Labeled Products. Bus Entrep Rev. 2016 Apr 13;14(1):15–32.
- Munekata PES, Pateiro M, Franco D, Lorenzo JM. Pork [Internet]. Boca Raton: CRC Press; 2021.
- Xie M, Lv X, Wang K, Zhou Y, Lin X. Advancements in Chemical and Biosensors for Point-of-Care Detection of Acrylamide. Sensors. 2024 May 29;24(11):3501.
- GODÁLOVÁ Z, BERGEROVÁ E, SIEKEL P. Effect of high temperature and pressure on quantification of MON 810 maize. Czech J Food Sci. 2013 Aug 31;31(4):376–381.
- Adenuga BM, Montowska M. A systematic review of DNA‐based methods in authentication of game and less common meat species. Compr Rev Food Sci Food Saf. 2023 May 29;22(3):2112–2160.
- Suzuki N, Uchikawa K, Yamada H, Chow S. Genetic Divergence and Identification of Two Controversial Lanternfishes (Actinopterygii: Myctophidae: Diaphus) Based on Mitochondrial Cytochrome b Sequences and PCR-RFLP Analysis. Species Divers. 2005;10(4):289–299.
- Bahuguna A, Singh A. Molecular characterization by using 12SrRNA and Cytochrome b for identification of species of genus Ratufa (Rodentia: Scuiridae) including Ratufa indica , endemic species of India. Mitochondrial DNA Part B. 2019 Jul 3;4(2):3085–3091.
- Feng C, Zhu X. Signal Amplification. In: Nano-Inspired Biosensors for Protein Assay with Clinical Applications. Elsevier; 2019. p. 287–312.
- Orbaniyah S, Hermawan A, Sudjadi S, Irnawati, Rohman A, Hastuti AAMB. The employment of species-specific primer in combination with real-time polymerase chain reaction for analysis of pork in meatball products. Food Res. 2024 Oct 30;8(5):438–442.
- Natonek-Wiśniewska M, Krzyścin P. Detection of the Species Composition of Food Using Mitochondrial DNA: Challenges and Possibilities of a Modern Laboratory. In: Biochemical Analysis Tools - Methods for Bio-Molecules Studies. IntechOpen; 2020.
- Anggita AD. Penggunaan Primer Mitochondrial Nadh Dehydrogenase Subunit 5 (Mt-Nd5) Sebagai Pendeteksi Cemaran Daging Babi Pada Bakso. Universitas Brawijaya; 2019.
- Tanabe S, Miyauchi E, Muneshige A, Mio K, Sato C, Sato M. PCR Method of Detecting Pork in Foods for Verifying Allergen Labeling and for Identifying Hidden Pork Ingredients in Processed Foods. Biosci Biotechnol Biochem. 2007 Jul 23;71(7):1663–1667.
- Friehs K. Plasmid Copy Number and Plasmid Stability. In 2004. p. 47–82.
- Li W, Olivier M. Current analysis platforms and methods for detecting copy number variation. Physiol Genomics. 2013 Jan;45(1):1–16.
- D’haene B, Vandesompele J, Hellemans J. Accurate and objective copy number profiling using real-time quantitative PCR. Methods. 2010 Apr;50(4):262–270.
- Elyasigorji Z, Izadpanah M, Hadi F, Zare M. Mitochondrial genes as strong molecular markers for species identification. Nucl. 2023 Apr 28;66(1):81–93.
- Yoshida T, Nomura T, Shinoda N, Kusuma T, Kadowaki K ichi, Sugiura K. Development of PCR primers for the detection of porcine DNA in feed using mtATP6 as the target sequence. J Food Hyg Soc Japan. 2009;50(2):89–92.
- Kusnadi J, Ashari N. The Sensitivity Evaluation of mt-DNA Genes; NADH Dehydrogenase Sub Unit 5 (ND5), D-Loop, and Cytochrome-b (Cty-b) to Detect Pork (Sus scrofa) DNA Isolate and DNA Fragment in Meatball using PCR Technique. In: Proceedings of The Food Ingredient Asia Conference (FiAC). SCITEPRESS - Science and Technology Publications; 2020. p. 12–19.