RESEARCH ARTICLE
Inhibition of Phospholipase A2 by Dihydrostilbene Glycoside Isolated from the Ethyl Acetate Fraction of Indigofera capitata: Structural Insights from IR and NMR
Academic Editor: Bharath Kumar Chagaleti
Sciences of Phytochemistry|Vol. 3, Issue 1, pp. 55-67 (2025)
CC BY 4.0-2025 Authors
Views
Downloads
Shares
Received
Jan 29, 2025Revised
Apr 8, 2025Accepted
May 1, 2025Published
May 19, 2025
Abstract
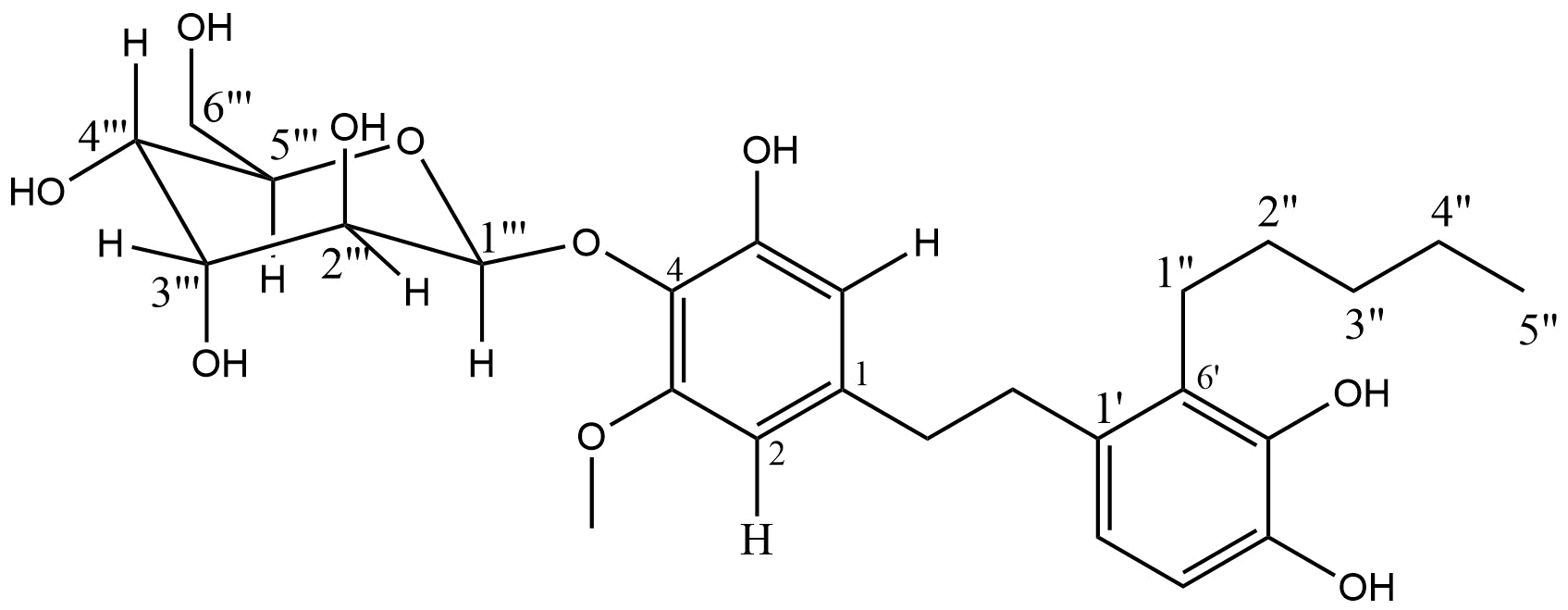
Snakebites and subsequent envenomations remain a significant health challenge, especially in rural regions of underdeveloped countries, where they contribute to high mortality rates. In Nigeria, particularly in rural areas, the scarcity of healthcare facilities and antivenoms exacerbates the issue. Rural communities commonly seek traditional plant-based remedies due to their affordability and cultural significance. This study investigates the ethyl acetate fraction of Indigofera capitata, a plant traditionally used to treat snakebites, for its potential to inhibit phospholipase A2 (PLA2) activity. Chromatographic separation of the ethyl acetate fraction led to the isolation of a greenish solid substance, coded MM2, identified as 3-methoxyl, 5, 4', 5' trihydroxyl, 6' pentyl-4-O-glycopyranoside through chromatographic and spectroscopic methods. The inhibition of PLA2 was assessed using an in vitro acidimetric assay. Compound MM2 was tested at 1 mg/mL, 0.1 mg/mL, and 0.01 mg/mL. The highest inhibition of 47.37% was observed at 1 mg/mL. At 0.1 mg/mL, the inhibition decreased to 36.84%, indicating reduced efficacy. Interestingly, at 0.01 mg/mL, the inhibition returned to 47.37%, suggesting that the enzyme inhibition by MM2 is concentration-independent within the tested range. The promising PLA2 inhibition observed with compound MM2 supports its potential as an antivenom agent. This research highlights the therapeutic potential of I. capitata in addressing the global snakebite crisis. To the best of our knowledge, this is the first report documenting this compound's isolation, characterization, and antivenin screening.
Introduction
Snakebites and their resulting envenomations remain a significant cause of mortality, particularly among rural populations in underdeveloped countries. This issue is classified as a class one neglected global health concern by the World Health Organization (WHO) (1). Although available epidemiological data on the incidence and fatalities due to snakebites are likely underestimated, this is primarily due to the lack of computerized and real-time research tools in regions such as Asia and Sub-Saharan Africa, where snakebite incidents are most prevalent (2-3). Additionally, the low reported incidence of snakebite injuries, especially in Africa, can be attributed to many victims not seeking medical care, which contradicts the WHO's recommended first aid guidelines for snakebite treatment in the region (4). Facilities are either severely inadequate or unavailable in many areas (4). In regions where facilities exist, essential medications like serum-based antivenoms, which require refrigeration, are often inaccessible due to unreliable power supplies. Given the critical time sensitivity for administering antivenom after a snakebite (before venom toxins cause significant harm), transporting patients to distant urban hospitals that might stock the necessary treatment is often ineffective (4-5). Moreover, conventional antivenoms are costly, making them unaffordable for many impoverished victims. In addition, these antivenoms can lead to serious side effects, such as anaphylactic shock, pyrogenic reactions, and serum sickness. They also fail to adequately address venom-induced complications like necrosis, hemorrhaging, cytotoxicity, and nephrotoxicity (5).
Cobras, originating from Africa and belonging to the Elapidae family, are the most commonly encountered snakes in hospital cases involving snakebites. These spitting cobras possess venom that causes ulceration, necrosis at the bite site, and systemic neurotoxic effects. Hospital data indicate that the black-necked spitting cobra (Naja nigricollis) is Northern Nigeria's most prevalent and clinically significant snake, especially in urban areas (6). A primary component of cobra venom is a high concentration of phospholipases A2 (PLA2), potent neurotoxins that specifically target receptors at presynaptic nerve terminals (7). This interaction results in the breakdown of phospholipids in the plasma membrane, disrupting the normal release of neurotransmitters and leading to hemolysis and hemorrhaging (8). The venom also contains L-amino acid oxidase, a flavoenzyme that promotes the oxidative deamination of L-amino acids, producing alpha-ketoacids, hydrogen peroxide (H2O2), and ammonia (7-9). These substances, particularly the free radical hydrogen peroxide, play a significant role in the pathophysiological effects and genotoxic consequences following envenomation. The clinical symptoms of cobra venom poisoning include bleeding, swelling, mild fever, anxiety, pain, difficulty swallowing and breathing, convulsions, and paralysis of the respiratory muscles and nerves, which can ultimately result in death (10-11).
In Nigeria, particularly in rural regions, healthcare facilities are often limited, and even when available, access to standard antivenoms is scarce (4, 6). According to Sani et al., the annual incidence of snakebites in the Savannah Region of Northern Nigeria is 497 per 100.000 people, with a mortality rate of 12.2% (12). A significant proportion of snakebite victims do not seek hospital treatment. In Northern Nigeria, it has been estimated that only 8.5% of snakebite victims go to hospitals. The low hospital attendance is primarily due to the belief that snakebites are not a condition requiring hospitalization, along with fears of death, superstition, amputation, and the high cost of antivenoms (4, 13).
For many years, plants have provided alternative methods for managing snakebites. Rural communities often turn to medicinal plants for disease treatment, valuing them for their safety, effectiveness, cultural significance, affordability, and easy availability (14-15). Over time, various efforts have been made to create antidotes for snake venom derived from plant sources. In many rural regions, traditional healers are typically the first to be consulted in snakebite cases, with some of these incidents successfully treated using folk remedies, particularly plant-based medicines (6, 16).
Indigofera capitata Kotschy (Fabaceae) is a shrub typically found in bushy areas and old farmlands, growing up to 1 meter in height across the savannah regions, from Senegal to Nigeria, and extending throughout Sudan, East Africa, Malawi, and Angola (17). It is locally known as "Baba" among the Hausa-speaking population in northern Nigeria. Traditionally, it has been used in folk medicine to treat jaundice, snakebites, and as a mouthwash (18). Previous pharmacological studies have shown that its methanol extract possesses antidiarrheal, antibacterial, and antivenom properties (19). As part of our ongoing search for bioactive compounds with antivenom potential from Nigerian plants, we report the isolation and characterization of a novel dihydrostilbene compound, 3-methoxyl, 5, 4', 5' trihydroxyl, 6' pentyl-4-O-glycopyranoside, derived from the aerial parts of I. capitata. The compound's structure was determined through chromatographic methods, IR, 1D and 2D NMR analysis, and comparison with existing literature data, and its inhibition of phospholipase A2 enzyme was evaluated in an animal model.
Experimental Section
Materials
Collection, Identification, and Preparation of Plant Material
The whole plant of I. capitata, growing in the wild, was harvested in September from the Basawa area of Zaria, Kaduna State, Nigeria. The plant was authenticated at the Herbarium section of the Department of Botany, Ahmadu Bello University, Zaria, by Mal. Namadi Sanusi, by comparing it with a herbarium voucher specimen (No. 470). The aerial parts were then cut, air-dried to a constant weight for 2 weeks, then size-reduced using a pestle and mortar into powdered material, weighed, and the powdered plant material was kept in a polythene bag for further use.
Chemicals
All organic solvents employed for extracting and fractionating plant material were of laboratory grade (Merck Millipore). The solvents utilized for column chromatography, the crystallization of compounds, and the chemicals used to assess antimalarial properties were of analytical grade. The standard chloroquine powder was purchased from Sigma Aldrich, St. Louis, MO, USA.
Source and Maintenance of Experimental Animals
Adult Swiss albino mice of either sex weighing 17–30 g were used. These were obtained from the Animal House Facility of the Department of Pharmacology and Therapeutics, Ahmadu Bello University, Zaria, Nigeria. The animals were fed on standard mice pellets, and water was supplied ad libitum. Before the tests, they were conditioned (22 ± 2 °C; 12: 12 h dark/light cycle), at ambient temperature, and given time to acclimatize for 2 weeks. All experimental protocols were approved by Ahmadu Bello University, Zaria, Committee on Animal Use and Care (ABUCAUC). The animals’ care and handling comply with the National Regulations for Animal Research.
Venom Sample
The venom of the N. nigricollis snake was collected by the usual milking method of Markfarlane (1967) (20) from a locally caught snake (kept at the herbarium, Department of Pharmacognosy and Drug Development, Ahmadu Bello University, Zaria, Nigeria). It was pooled, lyophilized, and stored at about 2-4 °C before the experiments. The venom was subsequently referred to as the crude venom of the snake, which was used as a standard to compare with the efficacy of the isolated compound.
Extraction and Fractionation
A total of 800 g of powdered I. capitata plant material was subjected to extraction using 2 L of methanol in a Soxhlet apparatus. The methanol extract was concentrated under reduced pressure, yielding 178 g of residue, designated as the I. capitata extract. From this crude methanol extract, 80 g was dissolved in distilled water and partitioned sequentially using various solvents: 1 L of n-hexane, 1 L of chloroform, 1.5 L of ethyl acetate, and 2 L of n-butanol. This process yielded the following fractions: n-hexane fraction (HF): 6.9 g, chloroform fraction (CF): 5.1 g, ethyl acetate fraction (EF): 6.5 g, and n-butanol fraction (BF): 22.1 g.
Isolation of Compound MM2 from Indigofera capitata
Column Chromatography of Ethyl Acetate Fraction
The comparative TLC analysis of the ethyl acetate fraction (EF) (5.1 g) and chloroform fraction (CF) (6.5 g) revealed similar major spots, prompting their combination into a single fraction, referred to as the ethyl acetate fraction (EF). This pooled fraction was then subjected to gel filtration chromatography, where it was eluted continuously with methanol. Thirty-eight collections containing 2-3 mL aliquots were made and pooled according to their TLC profiles, resulting in six fractions (A-F). Fraction B underwent further purification by silica column chromatography, using a continuous elution with a hexane: ethyl acetate solvent mixture in varying ratios (100: 0, 95: 5, 90: 10, 85: 15, etc.). Aliquots of 3-5 mL were collected, yielding 139 fractions (A1-K1). Fraction seven (G1) gave a single homogenous spot on TLC profile and was coded MM2.
Characterization of Isolated Compound MM2
The isolated compound was individually subjected to the following processes:
i. Physical appearance: The colour and physical appearance were observed and recorded.
ii. Chemical test: The isolated compound was subjected to the Liebermann-Buchard, Ferric chloride, and Fehling tests.
iii. Melting point determination: A melting point test was conducted on the isolated compound using a Gallenkamp electrothermal melting point apparatus. Results were uncorrected.
iv. Spectral analysis: Infrared (Agilent Technologies Cary 6030 FTIR spectrometer, Santa Clara, USA) and Nuclear Magnetic Resonance Spectroscopy (Bruker AVANCE III Instruments Incorporation, Billerica, MA, USA), 500 MHz and 125 MHz (1D and 2D) analyses were carried out on the isolated compound for characterization. Chemical shift values (δ) were reported in parts per million (ppm) relative to the appropriate internal solvent standard, and coupling constants (J values) are given in Hertz. HMBC spectra were optimized for a long-range JH-C of 7Hz (d6 = 0.07s). The NMR solvents used for these measurements are deuterated chloroform.
Acute Oral Toxicity Study
The LD99 of the venom was determined by the method described by Theakston and Reid (1983) (21). The N. nigricollis venom was reconstituted with normal saline, and concentrations ranging from 0.1 to 0.6 mg/mL were obtained. Thirty-five mice were divided into seven groups (n=5 mice each) and injected i.p. with different doses of the reconstituted venom. The control group received only normal saline (0.2 mL each i.p.). The LD99 was calculated using probit analysis (Finney, 1971) of death occurring within 24 h of venom injection. The LD99 of the venom was adopted from the reported works of Abubakar et al. (22-24) since they are from the same batch.
Antivenin Studies
The acidimetric assay for PLA2 enzymes was performed according to the method described by Tan and Tan (1988) (25). An equal volume of substrate consisting of calcium chloride (18 mM), sodium deoxycholate (8.1 mM), and egg yolk was mixed and stirred for 10 min to obtain a homogeneous suspension. The pH of the suspension was adjusted to 8.0 using 1 M sodium hydroxide. Snake venom (0.2 mg/mL) was added to the mixture (15 mL) to initiate hydrolysis, with DMSO as the control. A decrease in pH was measured after 2 min using a digital pH meter. A 1.0 unit drop in pH was equivalent to 133 µmoles of fatty acids released from the egg yolk mixture. The enzymatic activity of phospholipase A2 was calculated as the micromoles of fatty acids released per minute. The assay included a negative control (substrate without venom) to monitor spontaneous hydrolysis. The pH shifts in the test samples were corrected for any background activity in the negative control. The concentrations of MM2 (1 mg/mL, 0.1 mg/mL, and 0.01 mg/mL) were selected from isolation yields and previous research on plant-derived PLA2 inhibitors (22-24). Future studies will broaden this range to establish a full dose-response relationship.
To assess the antivenom potential of MM2 isolated from I. capitata, snake venom (0.2 mg) was pre-incubated with the isolated compound MM2 (at concentrations of 1.0, 0.1, and 0.01 mg/mL) to neutralize PLA2 hydrolytic activity. The inhibition of phospholipase activity by the compound was quantified and presented as percentage enzyme activity and inhibition.
Results
Isolation of MM2
Repeated Sephadex LH-20 column chromatography of the ethyl acetate fraction of the methanol extract yielded six pooled sub-fractions (Table 1). Sub-fraction B was subjected to silica gel column chromatographic separation, resulting in 139 collections pooled together based on TLC profile to give 11 fractions (A1-K1) (Table 2). Sub-fraction G1, coded MM2, appeared as a solid green substance and gave a single homogenous spot.
| S/No. | 2-3 mL aliquots | Codes |
|---|---|---|
| 1 | 1-7 | A |
| 2 | 8-14 | B |
| 3 | 15-17 | C |
| 4 | 18-22 | D |
| 5 | 23-28 | E |
| 6 | 29-38 | F |
| S/No | Solvent system (%) | Collections (3-5 mL) | Code |
|---|---|---|---|
| 1 | Hexane: EtOAc (100: 0) | 1-15 | A1 |
| 2 | Hexane: EtOAc (95: 5) | 16-50 | B1 |
| 3 | Hexane: EtOAc (90: 10) | 51-58 | C1 |
| 4 | Hexane: EtOAc (85: 5) | 59-67 | D1 |
| 5 | Hexane: EtOAc (80: 20) | 68-77 | E1 |
| 6 | Hexane: EtOAc (75: 25) | 78-89 | F1 |
| 7 | Hexane: EtOAc (70: 30) | 90-109 | G1 (MM2) |
| 8 | Hexane: EtOAc (65: 35) | 110-116 | H1 |
| 9 | Hexane: EtOAc (65: 35) | 117-123 | I1 |
| 10 | Hexane: EtOAc (60: 40) | 124-131 | J1 |
| 11 | Hexane: EtOAc (60: 40) | 132-139 | K1 |
Thin Layer Chromatographic Analysis of Compound MM2
Thin layer chromatographic (TLC) analysis of MM2 revealed a single homogeneous spot on TLC when developed with three different solvent systems (Table 3).
| Solvent systems | Rf value | Colour of spot after spray with 10% H2SO4 |
|---|---|---|
| Hexane: Ethyl acetate (7: 3) | 0.19 | Pink-violet |
| Hexane: Ethyl acetate (1: 1) | 0.62 | Pink-violet |
| Hexane: Ethyl acetate (1: 0.5) | 0.45 | Pink-violet |
Physicochemical Tests of Compound MM2
Solubility of Compound MM2
Compound MM2 was found to be soluble in chloroform and methanol.
Melting Point of Compound MM2
The compound was found to have an uncorrected melting point range of 135 - 137 °C.
Chemical Test of Compound MM2
When subjected to the ferric chloride test, it gave a blue/violet colouration.
Spectral Analysis of Compound MM2
FTIR Analysis of Compound MM2
The Fourier Transform Infra-red (IR) spectrum of compound MM2 showed absorption bands at 3380 cm-1 (moderate, br), 2926-2700 cm-1 (high), 1700-1509 cm-1 (high), 1457 cm-1 (high), 1375-1151 cm-1 (moderately high), 1088-700 cm-1 (moderate) as well as other weak bands (see Figure S1).
1H NMR Spectral Analysis
The 1H NMR spectrum (d ppm, 500MHz, CD3OD) of compound MM2 revealed the presence of the following major resonance values at dH 6.31 (1H, d, J=2.6 Hz, H-2), dH 6.39 (1H, d, J=2.6 Hz, H-6), dH 7.09 (1H, d, J=8.5 Hz, H-2ʹ), dH 6.69 (1H, d, J=8.5 Hz, H-3ʹ), dH 3.39 (2H, br, m, H-α), dH 2.77 (2H, br, m, H-β), dH 2.08 (2H, br, m, H-1ʹʹ), dH 2.35 (2H, br, m, H-2ʹʹ), dH 1.61 (2H, br, m, H-3ʹʹ), dH 1.32 (2H, br, m, H-4ʹʹ), dH 0.93 (3H, m, H-5ʹʹ), dH 3.67 (3H, s, 3-OCH3), dH 4.82 (1H, m, H-1ʹʹʹ), dH 3.50 (1H, m, H-2ʹʹʹ), dH 3.44 (1H, m, H-3ʹʹʹ), dH 3.43 (1H, m, H-4ʹʹʹ), dH 3.45 (1H, m, H-5ʹʹʹ), dH 3.75 (1H, m, H-6aʹʹʹ) and dH 3.89 (1H, m, H-6bʹʹʹ) (see Table 4; Figure S2).
| Position | 1H NMR (δH ppm) J in Hz | 13C NMR (δC ppm) | DEPT-135 |
|---|---|---|---|
| 1 | - | 114.1 | C |
| 2 | 6.31, 1H, d, J=2.6Hz | 111.1 | CH |
| 3 | - | 148.8 | C |
| 4 | - | 160.8 | C |
| 5, OH | - | 165.2 | C |
| 6 | 6.39, 1H, d, J=2.6Hz | 102.8 | CH |
| 1' | - | 135.5 | C |
| 2' | 7.09, 1H, d, J=8.5Hz | 130.7 | CH |
| 3' | 6.69, 1H, d, J=8.5Hz | 116.0, | CH |
| 4', 5', OH | - | 156.2 | C |
| 6' | - | 131.1 | C |
| Α | 3.39, br, m | 39.9 | CH2 |
| b | 2.77, br, m | 38.7 | CH2 |
| 1'' | 2.08, br, m | 28.3 | CH2 |
| 2'' | 2.35, br, m | 34.9 | CH2 |
| 3'' | 1.61, br, m | 26.2 | CH2 |
| 4'' | 1.32, br, s | 23.9 | CH2 |
| 5'' | 0.93, m | 14.5 | CH3 |
| 3-OCH3 | 3.67, s | 52.1 | -OCH3 |
| 1''' | 4.82 | 101.8 | CH |
| 2''' | 3.50 | 78.1 | CH |
| 3''' | 3.44 | 75.0 | CH |
| 4''' | 3.43 | 71.5 | CH |
| 5''' | 3.45 | 78.2 | CH |
| 6''' | 3.75, 3.89 | 62.5 | CH2 |
13C-NMR and 13C-DEPT NMR Spectral Analysis
The 13C-NMR (d ppm, 125 MHz, CD3OD) and DEPT 135 NMR experiments of compound MM2 indicated the presence of twenty-six (26) carbon peaks. Eight aromatic carbon peaks were observed for compound MM2 at dC 114.1 (C-1), dC 148.8 (C-3), dC 160.8 (C-4), dC 165.2 (C-5), dC 135.5 (C-1ʹ), dC 156.2 (C-4ʹ, 5ʹ) and dC 131.1 (C-6ʹ). Other peaks observed include; dC 111.1 (C-2), dC 102.8 (C-6), dC 130.7 (C-2ʹ), dC 116.0 (C-3ʹ), dC 39.9 (C-α), dC 38.7 (C-β), dC 28.3 (C-1ʹʹ), dC 34.9 (C-2ʹʹ), dC 26.2 (C-3ʹʹ), dC 23.9 (C-4ʹʹ), dC 14.5 (C-5ʹʹ), dC 52.1 (C-OCH3), dC 101.8 (C-1ʹʹʹ), dC 78.1 (C-2ʹʹʹ), dC 75.0 (C-3ʹʹʹ), dC 71.5 (C-4ʹʹʹ), dC 78.2 (C-5ʹʹʹ) and dC 62.5 (C-6ʹʹʹ) (see Table 4; Figure S3).
The DEPT-135 NMR experiments showed 8 quaternary, 9 methine, 7 methylene, 1 methoxy, and 1 methyl carbon peaks (Table 4; Figure S4).
HSQC Spectral Analysis
HSQC experiment of compound MM2 (CD3OD) establishes the attachment of various hydrogen/protons to their respective carbon atoms. The major correlations include; dH 6.31# dC 111.1, dH 6.39 # dC 102.8, dH 7.09 # dC 130.7, dH 6.69 # dC 116.0, dH 3.39 # dC 39.9, dH 2.77 # dC 38.7, dH 2.08 # dC 28.3, dH 2.35 # dC 34.9, dH 1.61 # dC 26.2, dH 1.32 # dC 23.9, dH 0.93 # dC 14.5, dH 3.67 # dC 52.1, dH 4.82 # dC 101.8, dH 3.50 # dC 78.1, dH 3.44 # dC 75.0, dH 3.43 # dC 71.5, dH 3.45 # dC 78.2 # dH 3.75 and dH 3.89 # dC 62.5 (see Table 4; Figure S5).
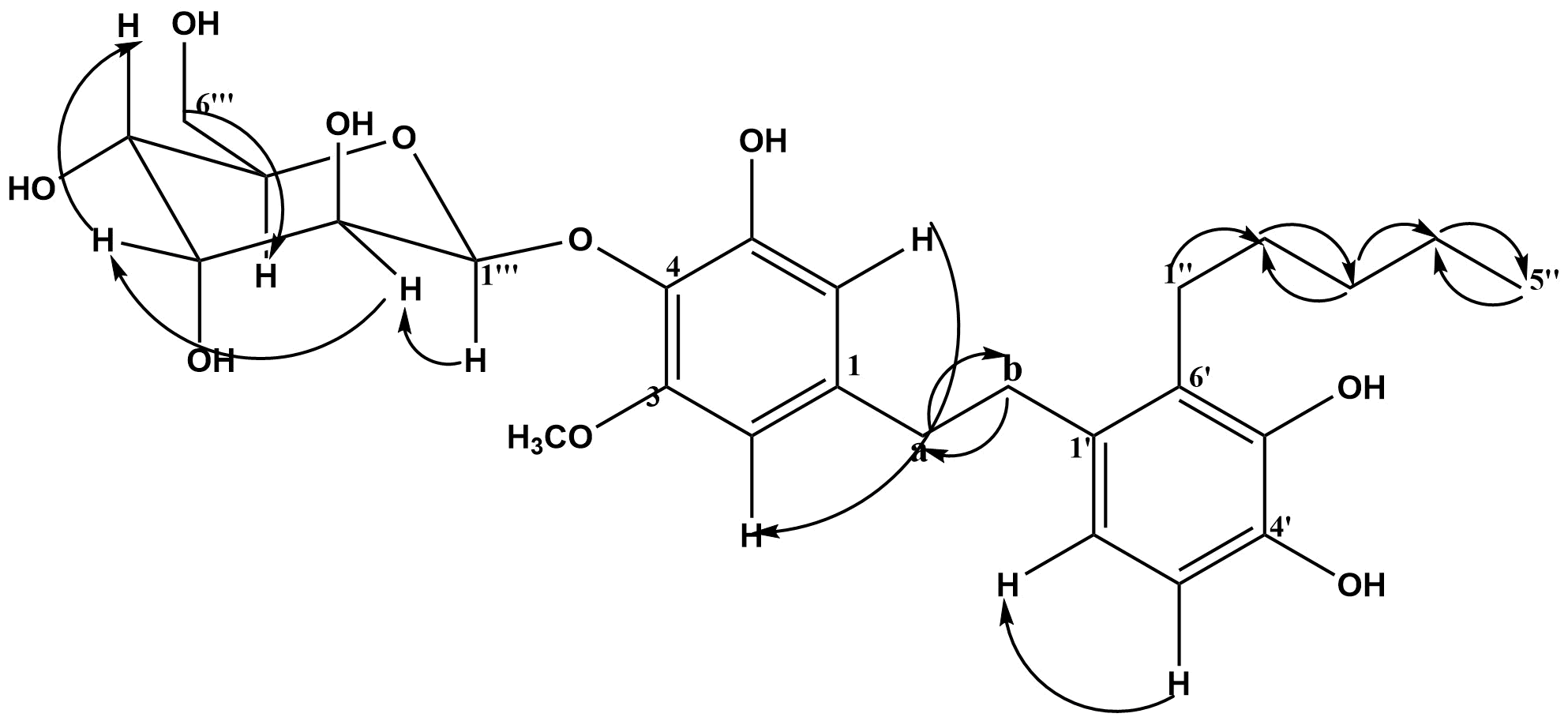
1H-1H COSY Spectral Analysis
The 1H-1H COSY spectra of compound MM2 established the correlation between the protons at dH 7.09 # dH 6.69, dH 6.39 # dH 6.31, dH 4.82 # dH 3.45, dH 3.89 # dH 3.75, dH 3.39 # dH 2.82, dH 2.35 # dH1.61, dH 2.08 # dH1.32, dH1.61 # dH1.32, dH1.32 # dH 0.93 (see Table 5; Figure S6).
| Protons | 1H-1H COSY | HMBC correlation | ||
|---|---|---|---|---|
| 2J (H→C) | 3J (H→C) | |||
| H-2 | H-6 | C-1 | C-4, C-6, C-α | |
| H-6 | H-2 | C-1, C-5 | C-4, C-2 | |
| H-2' | H-3ʹ | C-1', C-3' | C-b, C-6', C-4' | |
| H-3' | H-2ʹ | C-4' | C-1' | |
| H-α | H-β | C-1, C-b | C-3, C-1' | |
| H-b | H-α | C-α, C-1' | C-1, C-6', C-2' | |
| H-1'' | H-4ʹʹ | C-6', C-2'' | C-3', C-3'' | |
| H-2'' | H-3ʹʹ | C-1'', C-3'' | C-6', C-4'' | |
| H-3'' | H-2ʹʹ, 4ʹʹ | C-2'', C-4'' | C-1'', C-5'' | |
| H-4'' | H-1ʹʹ, 3ʹʹ, 5ʹʹ | C-3'', C-5'' | C-2'' | |
| H-5'' CH3O-3 H-1''' H-2''' H-3''' H-4''' H-5''' H-6''' | H-4ʹʹ H-2ʹʹʹ H-1ʹʹʹ - - H-6ʹʹʹ bH-5ʹʹʹ | C-4'' - C-2''' C-1''', C-3''' C-2''', C-4''' C-3''', C-5''' C-4''', C-6''' C-5''' | C-3'' C-3 C-3''', C-5''', C-4 C-4''' C-1''', C-5''' C-2''', C-6''' C-1''', C-3''' C-4''' | |
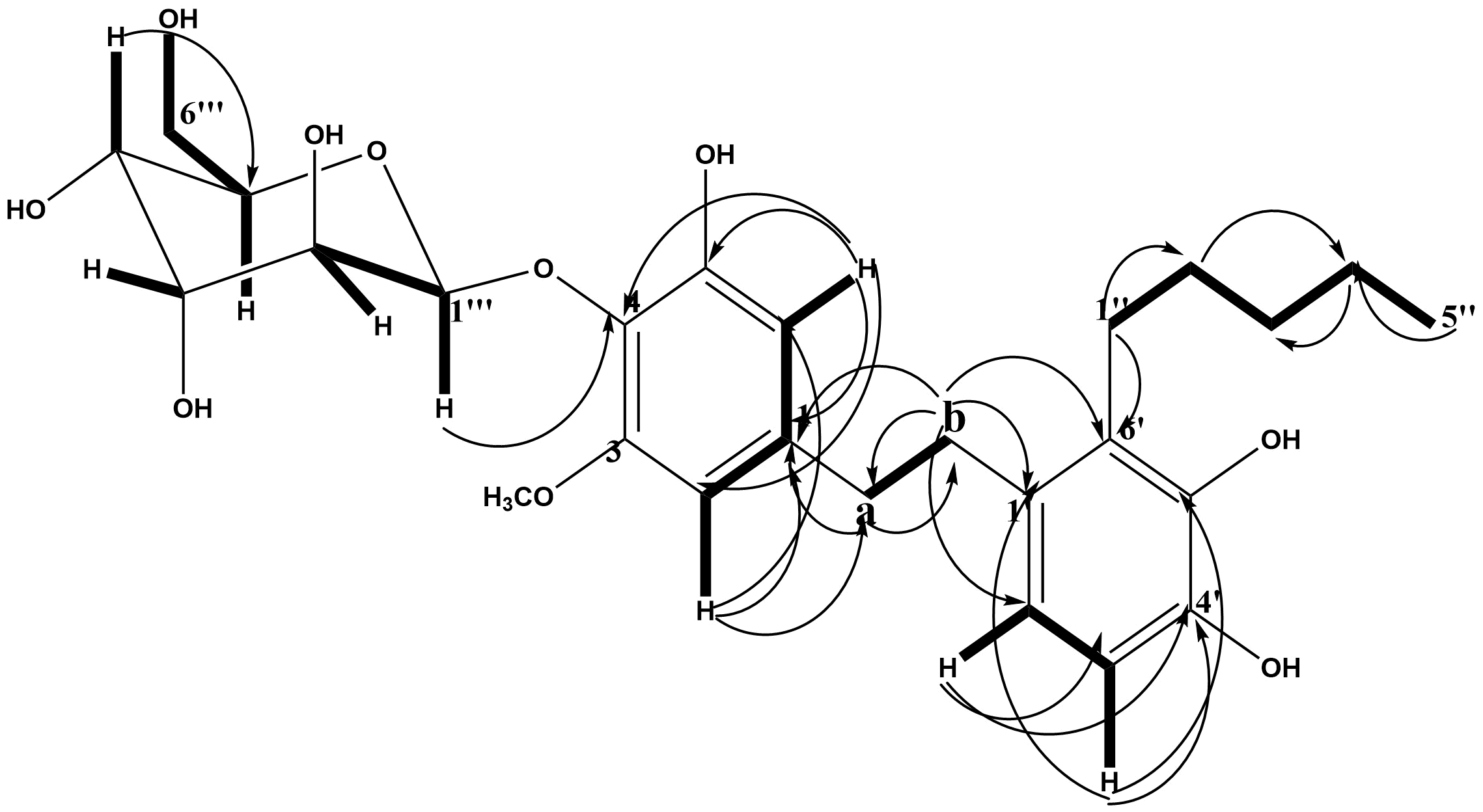
HMBC Spectral Analysis
The HMBC spectral analysis of compound MM2 established the connectivity between different fragments to the respective carbons with a distance range between two, three, and four (J2, 3-4) bonds apart within the molecule (see Table 5; Figure 2; Figure S7).
Pharmacological Studies
Median Lethal Dose LD99 of Naja Nigricollis Venom
The minimum lethal dose (LD99) of N. nigricollis venom was found to be 9.55 mg/kg.
Inhibition of Phospholipase A2 Enzyme by Compound MM2
The data presented in Table 6 suggest that compound MM2 exhibits enzyme inhibition against phospholipase A2 at various concentrations. Notably, at a dose of 1 mg/mL, compound MM2 showed the highest inhibition at 47.37%, which implies that at this concentration, the compound effectively reduces the enzyme activity. However, at a lower concentration of 0.1 mg/mL, the inhibition decreased to 36.84%, indicating a reduction in the compound's effectiveness as the dose was lowered. Interestingly, at an even lower concentration of 0.01 mg/mL, the inhibition was again 47.37%, which suggests that the enzyme inhibition by MM2 is independent of concentration within this range. This means that MM2 may reach a maximum inhibitory effect at doses as low as 0.01 mg/mL, and further increases in concentration do not seem to enhance its inhibition. This behavior indicates a concentration-independent inhibition, where the compound may either saturate the enzyme's active sites or undergo a mechanism where further increases in concentration do not yield proportionally higher inhibition. A 10% DMSO was used as a standard, which served as a control for the experimental setup.
| Doses (mg/mL) | % Enzyme inhibition |
|---|---|
| 1 | 47.37 |
| 0.1 | 36.84 |
| 0.01 | 47.37 |
Discussions
The isolation of compound MM2 was achieved through repeated gel filtration of the ethyl acetate fraction of the methanol extract, followed by silica gel column chromatography. The compound was obtained as a green solid with a melting point of 135-137 °C, indicating its high purity. Additionally, when tested with three different solvent systems, the compound exhibited a single spot on TLC, further confirming its purity and suggesting that it is a single compound (26).
The compound’s solubility in chloroform and methanol suggests it is moderately polar. Positive reactions with ferric chloride reagent indicate that the compound is likely phenolic (15, 27). Additionally, the pink-violet color observed upon spraying compound MM2 with 10% sulfuric acid and heating further supports a phenolic structure, potentially with a stilbenoid nucleus (15, 28).
The IR spectrum of compound MM2 (Figure S1) reveals a moderately intense broad band at 3380 cm⁻¹ and a moderate band at 1375 cm⁻¹, characteristic of O-H stretching vibrations, indicating the presence of an OH group. Alkyl C-H stretching is observed with a strong band at 2926 cm⁻¹ and a moderate band at 2855 cm⁻¹ (15). The absorption bands at 1718 cm⁻¹, 1595 cm⁻¹, and 1509 cm⁻¹, which are sharp and high-intensity, correspond to C=C double bonds. The IR absorptions at 3380 cm⁻¹ (O-H), 2926 cm⁻¹ (alkyl), 1595 cm⁻¹ (C=C aromatic), and 1375, 1237, 1203, 1151 cm⁻¹ (ether) support the presence of a phenolic structure. The methylene (CH₂)n vibrations are evident from a strong band at 1457 cm⁻¹. The moderate to high absorption at 1375-1151 cm⁻¹ is attributed to C-O vibrations, while the C-C vibration appears as a moderately intense band at 1032 cm⁻¹. Out-of-plane C-H vibrations from saturated and unsaturated components are observed in moderate bands between 1088-700 cm⁻¹ (29-31).
The 1H NMR spectrum (500MHz, CD3OD) exhibited signals for a methoxy group (δH 3.67), four (4) aromatic hydrogens; a meta-coupled (J=2.6Hz) aromatic hydrogen [(δH 6.31 (1H) and δH 6.39 (1H)], an ortho-coupled (J=8.5Hz) aromatic hydrogen [δH 7.09 (1H) and δH 6.69 (1H)]. There is a multiplet signal at δH 3.39-2.82, 4H, which was due to methylene hydrogens of the benzylic group that bridges between the two rings; this is a suggestion that the compound is a dihydrostilbene derivative (15, 28, 32).
In addition to these resonances, there is the presence of methyl hydrogens (δH 0.93, 3H, s) and other methylene hydrogens (δH 2.08, 2.35, 1.61, and 1.32). This indicated the presence of an aliphatic side chain in the molecule, suggesting that the compound has a side chain prenylation. Also, resonances at δH 4.82-3.43, br, m are attributed to a b- glycosyl unit linked to an aromatic ring. The signal at δH 4.82 (1H, br, s) was attributed to the presence of an anomeric proton.
The 13C and DEPT-135 NMR spectra (125 MHz, CD3OD) revealed signals for the methoxy, aromatic methines, benzylic methylene, oxygenated quaternary, quaternary, methylene, and other methine carbons. The assignment of these carbons, placement of methoxy, prenyl group, and the sugar moiety within the compound were achieved by the 2D experiments and reported literature (28).
In the HMBC experiment (Table 5), the meta coupled hydrogen H-2 (δH 6.31; dC 111.1 from HSQC) and H-6 (δH 6.39; dC 102.8 from HSQC) exhibited a common 2J correlation to an oxygenated quaternary carbon at dC 160.8 (C-4) and a 3J correlation to an aromatic quaternary carbon at 114.1 (C-1) from HMBC. The hydrogen at H-2 also has a common 2J correlation with another oxygenated quaternary carbon dC 148.8 (C-3). The methoxy group protons have common 3J connectivity to the carbon at dC 148.8 (C-3) from the HMBC, which is therefore attached to position 2. There is also a long-range correlation by the benzylic hydrogen H-α (δH 3.39; dC 39.9 from the HSQC) to the carbon at dC 148.8 (C-2) which confirms it further.
The hydrogen H-6 (δH 6.39; dC 102.8 from HSQC) in the HMBC showed a short-range correlation to an oxygenated quaternary carbon at dC 165.2 (C-5) from HMBC while a benzylic hydrogen H-α (δH 3.39; dC 39.9 from HSQC) showed a long-range correlation to the same carbon at dC 102.83 (C-6).
For the ortho-coupled hydrogens H-2' (δH 7.09; dC 130.7 from HSQC) and H-3', 5' (δH 6.69; dC 116.0 from HSQC), hydrogen H-2' has a 2J correlations to a carbon at dC 135.5 (C-1') and 116.0 (C-3') while the H-3' hydrogen has its common 2J correlations to a carbon at dC 130.7 (C-2'), dC 131.1 (C-6') and an oxygenated quaternary carbon at dC 156.2 (C-4'). There is a 3J correlation by the hydrogen H-2' to the benzylic methylene carbon at dC 38.7 (C-b), a carbon at dC 156.2 (C-4') and dC 131.1 (C-6'). These, together with the 2J correlation of hydrogen H-3' to carbon dC 131.1 and dC 156.2 allow the placement of carbon dC 156.2 as C-4'.
In the HMBC experiment, protons were attached to their respective carbons using the HSQC experiment. There was a 2J correlation between the methylene hydrogens H-1'' (δH 2.08; dC 28.3 from HSQC) of the prenylation chain to the quaternary carbon at 131.1 (C-6') of the aromatic ring (28, 33). The presence of an anomeric hydrogen at H-1''' (δH 4.82; dC 101.8 from the HSQC), confirmed the presence of the glucose moiety in the compound. The long-range correlation between the anomeric proton at δH 4.82 and carbon at (C 160.8 (C-4) confirmed that the glucose moiety is attached to position C-4.
Based on the physical properties, chemical test, spectral analysis, and literature (28, 33-34), the structure of the compound was proposed as 3-methoxyl, 5, 4', 5' trihydroxyl, 6'pentyl-4-O-glycopyranoside dihydrostilbene (see Figure 3).
Phospholipase A2 enzymes are abundant in snake venom and play a crucial role in developing toxic effects. These enzymes are responsible for several pathophysiological disorders, including cardiotoxicity, neurotoxicity, edema, hemolysis, and anticoagulation. As such, their contribution to harmful effects is undeniable, emphasizing the necessity for their inhibition (35).
In the acidimetric assay, PLA2 enzymes were hydrolyzed, releasing free fatty acids in sodium deoxycholate. The venom of N. nigricollis was shown to release free fatty acids, which were quantified by the decrease in pH of the egg yolk suspension. The isolated compound MM2 demonstrated the ability to inhibit the hydrolytic activity of phospholipase A2. The isolated compound MM2 displayed notable inhibition, with 47.4% activity at 1.0 mg/mL. Plant extracts have long been recognized as rich sources of pharmacologically bioactive compounds with antivenin properties (36). These compounds interact with toxins and enzymes in snake venom, inhibiting or neutralizing their actions. In several plant extracts, the active constituents responsible for antivenin effects have been identified, and their mechanisms of action have been suggested (37-39). Proteins and enzymes, including hyaluronidase, phospholipase, and hemorrhagins, are major components of snake venom (40). The toxic effects, such as lesions observed after N. nigricollis envenomation, are primarily due to these proteins and enzymes (40-42). The isolated compound MM2 protects against the toxic signs and lesions induced by envenomation. The non-linear inhibition profile of MM2 at tested concentrations suggests complex enzyme-compound interactions that could include saturation kinetics or allosteric effects. While these preliminary data indicate its inhibitory activity, a thorough dose-response study is needed to elucidate the mechanism and assess potency.
Naturally occurring substances, particularly plant polyphenols, are known to modify the activity of proteins and enzymes (43). Polyphenolic compounds, characterized by an aromatic ring with one or more hydroxyl groups and functional groups such as double bonds or oxygen-containing groups, can inhibit PLA2 activity (44). The inhibitory effects of the isolated compound MM2 may be attributed to these properties (45). The atypical dose-response of MM2 may result from aggregation, multi-site binding of PLA2, or interaction with detergent. Therefore, glycosidic stilbenes like MM2 aggregate into micellar formations at intermediate concentration, reducing bioaccessibility (44). Or, its pentyl side chain (see Table 4) might bind into PLA2 hydrophobic pockets, and saturation kinetics obviate classical dose-dependence. Such possibilities will be explored in future kinetic and biophysical work.
The inhibitory activity of MM2 (47.37%) is comparable to that of aristolochic acid (50%) and columbin (40%), two well-characterized plant-derived PLA2 inhibitors (22, 39). Though more potent, synthetic inhibitors like Varespladib are, MM2’s natural origin and structural novelty (e.g., glycosylation at C-4) may be more biocompatible. Further studies are needed to characterize its structure-activity relationship (SAR) with other polyphenolic inhibitors (44).
Conclusion
In this study, one compound was isolated from the ethyl acetate fraction of the I. capitata Kotschy plant. The results suggest that compound MM2 could be a potent inhibitor of phospholipase A2 at low concentrations, with its inhibitory effect reaching a plateau, making it an interesting compound for further investigation. However, the precise mechanism behind this concentration-independent inhibition would require additional studies to fully understand its mode of action. While MM2's non-linear inhibition profile (with comparable activity at 1 mg/mL and 0.01 mg/mL but reduced activity at 0.1 mg/mL) contravenes classical dose-response models, its structural features, a strikingly glycosylated dihydrostilbene scaffold and prenylated side chain, are suggestive of unique binding interactions with PLA2. Future studies will expand concentration ranges and employ biophysical tools to elucidate this behavior. To the best of our knowledge, this is the first report of the isolation of this compound from I. capitata and any other plant.
Abbreviations
1D = One Dimensional; 2D = Two Dimensional; BF = Butanol Fraction; CDCl3 = Deuterated Chloroform; Cf = Cloroform Fraction; COSY = Correlation Spectroscopic; 13C-NMR = Carbon-13 Nuclear Magnetic Resonance spectroscopy; DEPT = Distortionless Enhancement Polarization Transfer; EF = Ethyl acetate Fraction; FTIR = Fourier Transform Infrared Spectroscopy; HF = Hexane Fraction; 1H NMR = Proton nuclear magnetic resonance spectroscopy; HSQC = Heteronuclear Single Quantum Correlations; HMBC = Heteronuclear Multiple Bond Correlation; H2O2 = hydrogen peroxide; I = Indigofera; MeOD = Deuterated Methanol; MeOH = Methanol; PLA2 = phospholipases A2; TLC = Thin-layer chromatography; WHO = World Health Organization.
Declarations
Acknowledgment
We are thankful to all staff members of the Department of Chemistry, Kaduna State University and Department of Pharmaceutical and Medicinal Chemistry, Ahmadu Bello University Zaria, Kaduna. Nigeria.
Conflict of Interest
The authors declare no conflicting interest.
Data Availability
All data generated or analyzed during this study are included in this published article and its supplementary information files.
Ethics Statement
This study was conducted by the Declaration of Helsinki and approved by Ahmadu Bello University Zaria, Committee on Animal Use and Care (ABUCAUC). The collection of Indigofera capitata Kotschy plant (wild) used in the study complies with the national guidelines with no need for further affirmation. Indigofera capitata Kotschy plant (wild) was collected in bulk from the Basawa area of Zaria (Latitude: 11.13019, Longitude: 7.68581), Nigeria) in Kaduna, Nigeria. The plant was then identifed by Mal. Namadi Sanusi, and a voucher specimen was deposited at herbarium with ID (ABU470).
Funding Information
The authors declare that no financial support was received for the research, authorship, and/or publication of this article.
Supplemental Material
The supplemental data can be found at https://etflin.com/file/document/202501291541321374573495.docx. Figure S1. FTIR spectrum of compound MM2 of isolated from the ethyl acetate fraction Indigofera capitata Kotschy plant; Figure S2. Proton (1H) NMR Spectroscopy of compound MM2 isolated from ethyl acetate fraction of Indigofera capitata Kotschy plant; Figure S3. Proton decoupled 13C-NMR spectrum of MM2 isolated from ethyl acetate fraction of Indigofera capitata Kotschy plant; Figure S4. DEPT experiment of MM2 isolated from ethyl acetate fraction of Indigofera capitata Kotschy plant; Figure S5. HSQC NMR spectrum of MM2 isolated from ethyl acetate fraction of Indigofera capitata Kotschy plant; Figure S6. 1H-1H COSY spectrum of 19B isolated from ethyl acetate fraction of Indigofera capitata Kotschy plant; Figure S7. HMBC NMR spectrum of MM2 isolated from ethyl acetate fraction of Indigofera capitata Kotschy plant.
References
- Patra, A. and A.K. Mukherjee, 2021. Assessment of snakebite burdens, clinical features of envenomation, and strategies to improve snakebite management in Vietnam. Acta Trop., Vol. 216. 10.1016/j.actatropica.2021.105833.
- 2. Longbottom, J., F.M. Shearer, M. Devine, G. Alcoba and F. Chappuis et al., 2018. Vulnerability to snakebite envenoming: A global mapping of hotspots. Lancet, 392: 673-684.
- Sani, I., Ukwuani-Kwaja, A. N., Zubairu, A., Fakai, I. M., Bello, F., & Sani, H. A. Antivenom Potentials of Some Local Medicinal Plants Against Naja nigricollis Associated Envenoming in North-Western Nigeria.
- Chippaux, J.P., 2011. Estimate of the burden of snakebites in Sub-Saharan Africa: A meta-analytic approach. Toxicon, 57: 586-599
- Morais, V. (2018). Antivenom therapy: efficacy of premedication for the prevention of adverse reactions. J. Venom. Anim. Toxins Incl. Trop. Dis., 24:(7) 1-7
- Musah, Y., E.P.K. Ameade, D.K. Attuquayefio and L.H. Holbech, 2019. Epidemiology, ecology and human perceptions of snakebites in a savanna community of Northern Ghana. PLoS Negl. Trop. Dis., Vol. 13. 10.1371/journal.pntd.0007221
- World Health Organization, Guidelines For the Prevention and Clinical Management of Snakebite in Africa, 2010 accessed 29 june 2012
- Rossetto, O., Morbiato, L., Caccin, P., Rigoni, M., & Montecucco, C. (2006). Presynaptic enzymatic neurotoxins. Journal of neurochemistry, 97(6), 1534-1545.
- Adeyi, A. O., Ajisebiola, S. B., Adeyi, E. O., Alimba, C. G., & Okorie, U. G. (2020). Antivenom activity of Moringa oleifera leave against pathophysiological alterations, somatic mutation and biological activities of Naja nigricollis venom. Scientific African, 8, e00356.
- Lee, M. L., Fung, S. Y., Chung, I., Pailoor, J., Cheah, S. H., & Tan, N. H. (2014). King cobra (Ophiophagus hannah) venom L-amino acid oxidase induces apoptosis in PC-3 cells and suppresses PC-3 solid tumor growth in a tumor xenograft mouse model. International Journal of Medical Sciences, 11(6), 593.
- Marcussi, S., Stábeli, R. G., Santos-Filho, N. A., Menaldo, D. L., Pereira, L. L. S., Zuliani, J. P., ... & Soares, A. M. (2013). Genotoxic effect of Bothrops snake venoms and isolated toxins on human lymphocyte DNA. Toxicon, 65, 9-14.
- Sani, U.M., N.M. Jiya, P.K. Ibitoye and M.M. Ahmad, 2013. Presentation and outcome of snake bite among children in Sokoto, North-Western Nigeria. Sahel Med. J., 16: 148-153.
- Habib, A.G., 2013. Public health aspects of snakebite care in West Africa: Perspectives from Nigeria. J. Venomous Anim. Toxins Incl. Trop. Dis., Vol. 19. 10.1186/1678-9199-19-27.
- : Garba, D., Ali, B.H., Bawa, B., Sanusi, A., Sani, Y.M., Magaji, M.G., Abdullahi, M.I., Musa, A.M., Sadiya, H.H.. Exploring the Antimalarial Efficacy of Globimetula oreophila Leaf Fractions in Plasmodium berghei-Infected Mice: In Vivo Approach. Sciences of Phytochemistry. 2024; 3(2):105-113
- Garba, D., Ali, B. H., Bawa, B., Sanusi, A., Sani, Y. M., Magaji, M. G., ... & Sadiya, H. H. (2024). Isolation of secondary metabolites from leaves of Globimetula oreophila parasitizing Azadirachta indica: a spectroscopic study. Discover Plants, 1(1), 1-14.
- Haruna, A.K. and Choudhury, M.K (1995). In vivo antisnake venom actiivity of the furoid diterpene from Aristolochia albida Duch. Indian Journal of Pharmaceutical Science, 27: 222-224.
- Tamilselvi, N., R. Dhamotharan, P. Krishnamoorthy, and Shivakumar Shivakumar. "Anatomical studies of Indigofera aspalathoides Vahl (Fabaceae)." (2011): 738-746.
- Burkill, H.M. (1995). The Useful Plants of West Tropical Africa. Royal Botanic Garden Kew, U.K. 3: 361-670.
- Abdulkadir, S., Adamu, A.K, Dangora, D.B., Alonge, S.O., Ibrahim, G., Abubakar, M.S. and Yaro, A.H. (2007a). Phytochemical screening and antivenin activity of the methanolic extract of Indigofera capitata Kotchy in mice. Biological and Environmental Science Journal for the Tropics 4(2): 32-35.
- Markfarlane, R. G. (1967). Russel’s viper’s venoms, 1963-1964. British Journal of Haematology, 13: 437-451.
- Theakston, R.D.G. and Reid, H.A. (1983). Development of simple standard assay procedures for the characterization of snake venoms, ResearchBulletin of the World Health Organization, 61(6): 949-956
- Abubakar, M.S., Nok, A. J. and Balogun, E. (2001). Inhibition of Naja nigricollis Venom acidic Phospholipase A2 Catalyzed Hydrolysis of Ghost red blood cells by Columbin; Journal of Enzyme Inhibition and Medicinal Chemistry, 17(1): 55- 60.
- Isa, H.I., Ambali, S.F., Suleiman, M.M., Abubakar, M.S., Kawu, M.U., Shittu, M., Yusuf, P.O. and Habibu, B. (2015). In vitro neutralization of Naja nigricollis venom by stem-bark extracts of Commiphora africana A. Rich. (Burseraceae), Journal of Environmental Science, Toxicology and Food Technology,9(12): 100-105. e-ISSN: 2319-2402, www.iosrjournals.org
- Yusuf, A.J., Abdullahi, M.I., Haruna, A.K., Idris, A.Y. and Musa, A.M. (2015). Preliminary phytochemical screening, toxicological and antivenin property of the stem bark of Neocarya macrophylla on Naja nigricollis venom. African Journal of Pharmaceutical Research and Development, 7(1): 6-10.
- Tan, N. and Tan, C. (1988). Acidimetric Assay for Phospholipase A Using Egg Yolk Suspension as Substrate ’ Titration Curve of the Egg Yolk, 288: 282–288.
- John, D.R. (1964). Melting point tables of organic compounds (Utemark, Walter; Schicke, Walter). Journal of Chemical Education, 41(8): A590.
- Silva, G.L., Lee, I. and Douglas, K.A. (1998). Special problems with extraction of plants. In: Cannel, J. P. R. (ed.). Natural Products Isolation. Humana press publishers, New Jersey, USA. 356-358.
- Musa, A.M., Haruna, A.K., Ilyas, M., Ahmadu, A., Gibbons, S. and Rahman, M.M.(2008b). Dihydrostilbenes from Indigofera pulchra, Natural Product Communications, 3(5): 805-808.
- Kumar, R.S., Rajkapoor, B., Perumal, P. and Kumar, S.V. (2016). Beneficial effects of methanolic extract of Indigofera linnaei Ali. on the inflammatory and nociceptive responses in rodent models, Brazilian Journal of Pharmaceutical Sciences, 52(1):113 124,<http://dx.doi.org/10.1590/S1984- 82502016000100013
- Ramos-tejada, M.M. and Dura, J.D.G. (2002). Investigation of alumina / ( + ) -catechin system properties . Part I : a study of the system by FTIR-UV – Vis spectroscopy, Colloids and Surfaces Biointerfaces,24: 297–308.
- SenthilKumar, R., Rajkapoor, B., Perumal, P., Sekar V.K. and Suba Geetha, A. (2016). Beneficial effects of methanolic extract of Indigofera linnaei Ali. on the inflammatoryand nociceptive responses in rodent models, Brazilian Journal of Pharmaceutical Sciences, 52(1): 113-123. <http://dx.doi.org/10.1590/S1984-82502016000100013
- Zhao, W-M., Qin, G-W., Ye, Y., Xu, R-S. and Le, X-F. (1995). Benzyls from Stemona tuberosa. Phytochemistry, 38: 711-713.
- Aisyah, S.K., Norizan, A., Nur, F.A. and Khong, H.Y. (2018). Prenylated Dihydrostilbenes from Macaranga heynei (Euphorbiaceae), MalaysianJournal of Analytical Sciences, 22(2): 258-263. ISSN: 1394-2506
- Ersam, T., Fatmawati, S., and Fauzia, D.N. (2016). New prenylated stilbenes and antioxidant activities of Cajanus cajan (L.) millsp. (Pigeon pea), (September). https://doi.org/10.14499/ijc-v16i2p151-155.
- Asad, M.H.H.B., Sabih Darr-e-, Yaqab Tahir, Murtaza, G., Hussain M.S., Nasir, M.T., Azhar S., Khan S.A. and Hussain I. (2014). Phospholipases A2: Enzymatic assay for snake venom (Naja Naja karachiensis) with their neutralization by medicinal plants of Pakistan, Natural Drugs Research, 71(4): 625-630. ISSN: 0001-6837
- Gomes, A., Das, R., Sarkhel, S., Mishra, R., Mukherjee, S., Bhattacharya, S. and Gomes, A. (2010). Herbs and herbal constituents active against snake bite, Indian Journal of Experimental Biology, 48: 865-878. Review Article.
- Bhat, M.K., Kasturi, S. and Gowda, T.V. (1987). Structure-function relationship among neurotoxic phospholipase: NN-XIII-PLA2 from Indian cobra (Naja nigricollis) and VRVPL-V from Russell’s viper (Viper russelli) venoms. Journal of the International Society onToxicology, 29(1): 97-105.
- Tsai, L.H., Yang, L.L. and Chang, C. (1980). Inactivation of Formosan snake venoms in vivo by Aristolochic acid the chemical components of Aristolocia radix. Tai-Wan ko’Hsueh. 34(2): 40-44 Taiwanese.
- Vishwanath, B.S., Appu Rao, A.G. and Gowda, T.V. (1987). Interaction of phospholipase A2 from Vipera russelli venom with Aristolochic acid: a circular dichroism study, Journal of the International Society onToxicology,25: 939- 946.
- Warrell, D.A., Greenwood, B.M., Davidson, N.M., Omerod, L.D. and Prentice, C.R. (1976). “Necrosis, haemorrhage and complement depletion following bites by the spitting cobra (Naja nigricollis),” Quarterly Journal of Medicine, 45(177): 1-22.
- Warrell, D.A. (1987). Venomous bites and stings. In Bell Manson's Tropical Diseases, Mason Bahr PEC (ed.). Ballier Tindall: London; 855-898.
- Pugh, R.N.H and Theakston, R.D.G (1980). Incidences and mortality of snakebite in savannah Nigeria. Lancet, 11(29): 1181-1183.
- Haslam, E., Lilley, T.H., Cai, Y., Martin, R. and Magnolato, D. (1989). Traditional herbal medicines- the role of pholyphenols. Planta Medica, 55(1): 1- 8.
- Lindahl, M. and Tagesson, C. (1997). Flavonoids as phospholipase A2 inhibitors: importance of their structure for selective inhibition of group II phospholipase A2. Inflammation, 21(3): 347-356.
- Yusuf, A.J., Abdullahi, M.I., Haruna, A.K., Idris, A.Y. and Musa, A.M. (2015). Preliminary phytochemical screening, toxicological and antivenin property of the stem bark of Neocarya macrophylla on Naja nigricollis venom. African Journal of Pharmaceutical Research and Development, 7(1): 6-10.